-->
1. Gagasan awal tentang sel muncul pada abad XVII ketika seorang ahli memeriksa gabus di bawah kaca pembesar. Ahli tersebut adalah . . . .
1. Gagasan awal tentang sel muncul pada abad XVII ketika seorang ahli memeriksa gabus di bawah kaca pembesar. Ahli tersebut adalah . . . .
A. Robert Hooke
B. Theodor Schwann
C. Matthias Jokob Schleiden
D. Antonie Van Leuwenhoek
E. Carolus Linnaeus
2. Yang merupakan perbedaan sel hewan dan sel tumbuhan adalah . . . .
A. pada hewan terdapat plastida pada tumbuhan tidak ada
B. pada hewan tidak terdapat badan golgi, pada tumbuhan ada badan golgi
C. pada hewan terdapat badan golgi, pada tumbuhan ada badan golgi
D. pada tumbuhan tidak terdapat mikrotubulus pada hewan ada
E. pada hewan ada vakuola yang besar, pada tumbuhan ada vakuola tetapi kecil
3. Organel yang berperan dalam respirasi sel adalah . . . .
A. nukleus
B. retikulum endoplasma
C. ribosom
D. lisosom
E. mitokondria
4. Komponen sel yang tidak ditemukan pada sel hewan adalah . . . .
A. ribosom
B. badan golgi
C. plastida
D. mitokondria
E. sentrosom
5. Transpor zat melalui membran plasma dapat berlangsung dengan transpor aktif, transpor aktif adalah . . . .
A. transpor yang memerlukan energi untuk keluar masuknya molekul zat melalui membran
B. transpor dari larutan yang konsentrasinya rendah ke larutan yang konsentrasinya tinggi
C. transpor dari larutan yang konsentrasinya tinggi ke larutan yang konsentrasinya rendah
D. transpor yang mengendalikan pertukaran zat tanpa memerlukan energi
E. transpor yang dipengaruhi oleh ion natrium tanpa memerlukan energy
6. Beberapa jenis sel dapat memasukkan zat-zat padat atau tetes-tetes cairan melalui membran sel peristiwa semacam ini dinamakan . . . .
A. eksositosis
B. endositosis
C. difusi
D. osmosis
E. plasmolisis
7. Suatu selaput yang membungkus suatu massa protoplasma dinamakan . . . .
A. sitoplasma
B. nukleoplasma
C. mitokondria
D. retikulum endoplasma
E. membran plasma
8. Di dalam nukleus, gen terdapat di dalam . . . .
A. nukleolus
B. nukleoplasma
C. karioteka
D. kromatin
E. sitoplasma
9. Fungsi ribosom adalah . . . .
A. ekskresi sel
B. proses respirasi sel
C. tempat sintesis protein
D. organ pencerna intraselular
E. menyintesis lemak dan pati
10. Leukoplas yang berfungsi untuk membentuk dan menyimpan lemak adalah . . . .
A. amiloplas
B. elaioplas
C. kloroplas
D. kromoplas
E. plastid
11. Jaringan yang terdapat pada ujung-ujung tanaman (titik tumbuh) adalah . . . .
A. jaringan epidermis
B. jaringan endodermis
C. jaringan parenkim
D. jaringan penunjang
E. jaringan meristem
12. Aktivitas meristem primer akan mengakibatkan . . . .
A. bertambah panjangnya akar dan batang
B. membesarnya akar dan batang
C. terbentuknya pembuluh kayu
D. terbentuknya pembuluh kulit kayu
E. terbentuknya xilem dan floem
13. Beberapa jaringan pada tumbuhan adalah . . . .
- Epidermis
- Kambium
- Xilem
- Palisade
Jaringan yang hanya terdapat pada daun adalah . . . .
A. 1, 2, dan 3
B. 1, 3, dan 4
C. 1, 3, dan 5
D. 1, 4, dan 5
E. 2, 3, dan 5
14. Yang bukan jaringan penyusun akar tanaman monokotil adalah . . . .
A. epidermis
B. kambium ikatan pembuluh
C. korteks
D. perisikel
E. endodermis
15. Yang merupakan jaringan dewasa adalah . . . .
A. felogen
B. histogen
C. meristem primer
D. meristem apikal
E. meristem sekunder
16. Terbentuknya lingkaran tahun merupakan hasil aktivitas jaringan . . . .
A. kambium
B. meristem
C. histogen
D. felogen
E. dermatogens
17. Pembentukan akar cabang pada tumbuhan dikotil terjadi karena aktivitas . . . .
A. floem
B. endodermis
C. parenkim
D. perisikel
E. korteks
18. Fungsi kambium intravaskuler adalah . . . .
A. ke luar membuat floem, ke dalam membuat xilem
B. ke luar membuat korteks, ke dalam membuat kayu
C. ke luar membuat kulit, ke dalam membuat kayu
D. ke luar membuat korteks, ke dalam membuat xilem
E. ke luar membuat xilem, ke dalam membuat floem
19. Pernyataan berikut merupakan ciri jaringan epidermis, kecuali . . . .
A. sel-selnya berbentuk seperti balok
B. dilengkapi lapisan lilin
C. bermodifikasi menjadi stomata
D. mengandung banyak kloroplas
E. sel-selnya tersusun rapat
20. Sel pengiring terdapat pada jaringan . . . .
A. xilem
B. parenkim
C. floem
D. empulur
E. jari-jari empulur
21. Jaringan embrional yang terdiri atas tiga lapis disebut triploblastik. Lapisan jaringan tersebut urut dari luar ke dalam adalah . . . .
A. mesoderm - ektoderm - entoderm
B. ektoderm - mesoglea - entoderm
C. ektoderm - mesoderm - entoderm
D. entoderm - mesoderm - ektoderm
E. mesoglea - mesoderm - entoderm
22.Epitel yang melapisi kapiler darah, glomerulus, nefron dan alveolus paru-paru berbentuk . . . .
A. kubus berlapis tunggal
B. silindris berlapis banyak
C. kubus berlapis banyak
D. pipih berlapis banyak
E. pipih berlapis tunggal
23. Berikut adalah fungsi jaringan epitel, kecuali . . . .
A. pelindung atau proteksi
B. penghasil getah
C. penerima rangsang
D. pintu gerbang zat
E. pengangkut zat
24. Sel-sel penyusun sistem saraf disebut . . . .
A. neurolema
B. dendrit
C. akson
D. neuron
E. neurit
25. Sifat serat otot polos adalah . . . .
A. berinti satu yang terletak di tengah
B. membentuk otot jantung
C. bekerja di bawah kesadaran
D. mempunyai benang-benang fibril yang mempunyai garis terang dan garis gelap
E. dapat berkontraksi dengan kuat
26. Yang bukan merupakan jaringan penyokong adalah . . . .
A. darah
B. jaringan ikat
C. jaringan otot
D. jaringan tulang rawan
E. jaringan tulang
27. Jaringan tulang rawan terdapat pada . . . .
A. batang tenggorok
B. permukaan tulang rusuk
C. ujung-ujung tulang rusuk
D. daun telinga
E. tulang pipa
28. Sel darah putih yang bergranula terdiri atas . . . .
A. limfosit, neutrofil, dan eosinofil
B. neutrofil, eosinofil, dan basofil
C. eosinofil, eritrosit, dan monosit
D. eosinofil, basofil, dan monosit
E. neutrofil, basofil, dan limfosit
29.Di antara organ-organ berikut yang saling berinteraksi menyusun sistem pencernaan adalah . . . .
A. hati-usus-ginjal-limfa
B. pankreas-usus-hati-otot
C. lambung-limfa-paru-paru-usus-jantung
D. usus-lambung-hati-pankreas
E. jantung-paru-paru-usus-jantung
30.Sistem organ pada tubuh manusia yang bertanggung jawab untuk memindahkan hasil-hasil metabolisme yang tidak berguna ke luar dari tubuh dan menjaga keseimbangan sel dengan lingkungannya adalah . . . .
A. sistem pencernaan
B. sistem peredaran
C. sistem pernapasan
D. sistem ekskresi
E. sistem reproduksi
31. Otot disebut alat gerak aktif, sebab otot . . . .
A. melekat pada otot rangka
B. mempunyai kemampuan berkontraksi
C. tersusun atas senyawa protein rangkap
D. menghubungkan tulang dengan kulit
E. mempunyai plasma otot
32. Jumlah ruas tulang kelangkang manusia adalah . . . .
A. 4 ruas
B. 5 ruas
C. 6 ruas
D. 7 ruas
E. 12 ruas
33. Sendi putar adalah bentuk sendi yang memungkinkan gerakan . . . .
A. bebas dan berporos tiga
B. rotasi dan berporos satu
C. maju-mundur dan berporos dua
D. menggeser dan tak berporos
E. terbatas dan tak berporos
34. Berikut adalah fungsi rangka pada vertebrata, kecuali . . . .
A. membentuk sel darah
B. melindungi alat tubuh yang lemah
C. alat gerak pasif
D. menunjang tegaknya tubuh
E. tempat melekatnya otot polos
35. Pembengkokan tulang belakang ke arah samping disebut . . . .
A. lordosis
B. kifosis
C. skoliosis
D. layuh semu
E. fraktura
36. Kram atau kejang otot dapat terjadi karena . . . .
A. otot keras, kaya asam laktat, dan suhu dingin
B. tulang retak, otot lelah, dan suhu panas
C. sendi tulang infeksi, dingin, dan cukup energi
D. otot dan tulang melekat, cukup energi dan dingin
E. infeksi sendi, suhu dingin, dan cukup energi
37. Perlekatan otot pada tulang yang tidak bisa digerakkan disebut . . . .
A. insersio
B. sliding filamen
C. origo
D. pronasi
E. supinasi
38. Osteoporosis merupakan penyakit pada tulang yaitu . . . .
A. infeksi sendi
B. memar
C. keropos tulang
D. patah tulang
E. retak tulang
39. Energi bagi otot adalah . . . .
A. ADP
B. ATP
C. DNA
D. RNA
E. Aktin
40.Penyakit yang menyebabkan otot menjadi kejang karena toksin dari bakteri Clostridium tetani, yaitu . . . .
A. atrofi
B. hipertrofi
C. hernia abdominal
D. distrofi otot








 Pengertian
Pengertian
Lisosom ditemukan oleh de Duve dkk., pada awal tahun 1950 setelah mempelajari distribusi beberapa jenis enzim yang terlibat di dalam metabolisme karbohidrat. Salah satu enzim yang terlibat di dalam metabolisme karbohidrat. Salah satu enzim yang dipelajari adalah fosfatase asam yang memecah gugus fosfat pada beberapa fosfat yang mengandung ester fosfat.
kelompok lisosom dekat mitokondria
(Sheeler dan Bianchii, 1983)
Lisosom adalah vesikula yang berbatas membran dimana di dalamnya terkandung enzim-enzim hidrolase. Suatu organel dapat didefinisikan sebagai lisosom bilamana memenuhi beberapa kriteria, yaitu:
1. Organel yang bersangkutan berbatas membran;
2. Mengandung dua atau lebih enzim-enzim hidrolase yang semuanya adalah asam hidrolase;
3. Memiliki sifat kelatenan enzim.
Lisosom dibatasi oleh membran tunggal dan di dalamnya terkandung kurang lebih 40 jenis enzim yang semuanya adalah enzim hidrolase seperti protease, nuklease, glikosidase, lipase, fosfolipase, fosfatase, dan sulfatase. Enzim-enzim hidrolase bekerja dengan baik pada pH kurang lebih 5. Untuk mempertahankan pH tersebut, maka secara terus menerus terjadi pemompaan ion hidrogen ke dalam lumen lisosom dengan melibatkan hidrolisis ATP sebagai sumber energi. Lisosom memiliki protein maker yang disebut sebagai “Docking-marker acceptor”. Dengan demikian, lisosom akan dapat berfusi dengan vesikula-vesikula target dengan tepat.
Beberapa molekul sederhana dapat menembus membran lisosom, misalnya quinakrin. Quinakrin dapat meningkatkan pH di dalam lisosom jika diberikan ke dalam sel. Quinakrin digunakan di laboratorium sebagai inhibitor fungsi lisosom. Lisosom memiliki fungi pencernaan intra sel yang sangat luas meliputi pencernaan bahan-bahan intra dan ekstra sel, mikroorganisme yang telah difagositosis dan program kematian sel selama organogenesis oleh sel fagosit (Thorpe, 1984)
Enzim-enzim hidrolase memiliki kemampuan untuk memecahkan berbagai jenis molekul organik seperti polisakarida, protein, lipida, fosfolipida, dan asam-asam nukleat.
B. Pembentukan dan Nasib Lisosom
Asal dan pembentukan lisosom telah dipelajari dengan sangat intensif. Dari berbagai hasil temuan, ada dua pendapat yang berkenaan dengan asal dan pembentukan lisosom, yaitu:
1. Berbagai bukti telah ditemukan bahwa protein-protein hidrolitik dibentuk oleh ribsom yang terdapat pada retikulum endoplasma. Dari retikulum endoplasma kasar, selanjutnya protein tersebut ditranslokasikan menuju permukaan pembentukan badan golgi untuk diproses lebih lanjut. Setelah itu, protein-protein hidrolitik dikemas dan dibungkus dalam bentuk vesikula-vesikula untuk selanjutnya dilepaskan sebagai lisosom primer.
2. Protein-protein hidrolitik dibentuk pada ribosom yang terdapat pada retikulum endoplasma kasar, selanjutnya ia dilepaskan dalam bentuk vesikula menuju daerah GERL (Golgi associated Endoplasmic Reticulum giving rise to Lisosom) yang berdekatan dengan daerah permukaan matang badan golgi. Dari GERL, selanjutnya dilepaskan vesiula-vesikula yang disebut lisosom primer .
Gambar 7.4 Dua ide tentang pembentukan
lisosom (Thorpe, 1984)
Lisosom yang pertama dibentuk oleh sel dan belum terlibat dalam aktivitas pencernaan sel disebut lisosom primer. Sedangkan lisosom sekunder adalah lisosom yang merupakan hasil fusi berulang antara lisosom primer dengan berbagai substrat yang berbatas membran (Albert et al., 1983). Dengan demikian, lisosom sekunder telah terlibat dalam aktivitas pencernaan sel dan di dalam lumennya terdapat substrat dan enzim-enzim hidrolitik. Lisosom sekunder memiliki dua fungsi yang berbeda, yaitu:
1. Heterolisosom, yaitu bila substrat yang dicerna berasal dari luar sel. Dengan demikian, heterolisosom dibentuk dari hasil fusi antara lisosom primer dengan fagosom atau endosom. Heterolisosom sering disebut sebagai vakuola pencerna.
Albert et al. (1983) membagi heterolisosom menjadi dua tipe, yaitu:
Di dalam sel, sesungguhnya terdapat kerjasama yang erat antara heterolisosom dengan autolisosom. Selama heterofagi berlangsung, proteinprotein ditempatkan di dalam vesikula-vesikula endosom, kemudian berfusi dengan lisosom primer dan selanjutnya mengalami hidrolisis. Selama autofagi, sitosegresom berfusi dengan lisosom primer membentuk autolisosom dan memasuki siklus pencernaan intrasel (gambar 7.6)
Tergantung pada keadaan fisiologisnya, vakuola pencerna atau vakuola autofagi pada akhirnya mengalami satu dari tiga kemungkinan yang terjadi, yaitu :
1. Mengosongkan kandungannya dengan cara eksositosis atau defekasi seluler;
2. Menjadi bahan residu tanpa bahan hidrolase;
3. Menghidrolisis kandungannya secara sempurna untuk dapat berdifusi dan selanjutnya siap untuk siklus aktivitas yang baru.
Gambar 7.7. Nasib lisosom (Thorpe, 1984)
Tentang pembentukan sitosegresom, ada beberapa pandangan yang diusulkan, yaitu :
1. Sitosegresom dibentuk dari suatu membran sisterna yang melingkupi mitokondria secara sempurna dan selanjutnya diikuti dengan berdegenerasinya membran dalam;
2. Sitosegresom dibentuk dari suatu membran yang melingkupi mitokondria secara sempurna
3. Sitosegresom dibentuk dari vesikula endosom yang melingkupi mitokondria secara sempurna yang selanjutnya diikuti dengan berdegenerasinya membran dalam;
4. Sitosegresom dibentuk dari vesikula endosom, dimana mitokondria memasuki vesikula endosom melalui suatu celah.
Bentuk akhir heteroslisosom dan autolisoson disebut telolisosom atau postlisosom atau badan residu. Bahan-bahan yang terkandung di dalam telolisosom sewaktu-waktu dapat dilepaskan. Proses pelepasannya dinamakan defekasi seluler. Bahan-bahan yang telah dicerna di dalam lisosom dapat kembali dilepaskan ke dalam sitoplasma dan selanjutnya terlibat di dalam proses katabolisme atau anabolisme.
C. Ultra Struktur dan Komposisi Lisosom
Lisosom merupakan vesikula berbatas membran tunggal, dimana di dalamnya terdapat enzim-enzim proteolitik. Membran lisosom mengandung karbohidrat netral, hexoamina, dan asan N-asetilmuramat yang lebih banyak dibandingkan dengan membran plasma.
Pengamatan dengan mikroskop elektron menunjukkan bahwa membran lisosom memiliki membran dengan tebal 9 nm, lebih tebal dari membran mitokondria. Membran lisosom memiliki kemampuan untuk berfusi secara selektif dengan membran sel yang lain, seperti fusi yang terjadi antara membran lisosom dengan fagosom atau endosom selama pencernaan intra sel. Demikian pula antara lisosom dengan membran plasma selama berlangsungnya sekresi sel.
Peranan Lisosom
Peranan fisiologi lisosom umumnya berhubungan dengan pencernaan intraseluler. Misalnya pencernaan makanan yang berlangsung pada protozoa dimana bahan-bahan yang berasal dari luar dicerna secara intraseluler atau heterofagi.
Endositosis merupakan persyaratan bagi pencernaan intraseluler bahan eksogen dengan molekul tinggi. Bukti menunjukkan bahwa vakuola makanan dihasilkan dari fusi antara endosom dengan lisosom primer.
Dalam darah, terdapat banyak sel-sel fagosit yang bekerja sebagai penghalang yang efektif dari invasi mikroorganisme atau benda-benda asing lainnya. Ada empat tipe fagosit darah, yaitu (i) Polymorpho Nuclear Neutrofic Leukocytes (PMNs), (ii) Eosinofil, (iii) Basofil, dan (iv) Monosit. Meskipun keempat tipe tersebut bersirkulasi di dalam darah, neutrofil dan manosit memiliki kemampuan untuk meninggalkan aliran darah dan mengembara di seluruh jaringan untuk menghilangkan bahan-bahan asing dalam jaringan dengan cara memfagositosisnya.
Monosit di dalam jaringan akan berkembang menjadi dewasa dan menjadi sel-sel makrofag. Berbagai jenis makrofag antara lain histiosit dalam jaringan pengikat, makrofag alveolar di dalam paru-paru, sel-sel kuffer di dalam jaringan hati, makrofag pelural di dalam peritoneal, osteoklas di dalam tulang, sel mikroglia di dalam sistim saraf pusat, sel schwann di dalam serabut saraf perifer, sel sinvial tipe A di dalam ruang sendi, dan makrofag di dalam jaringan limfoid dan jaringan ikat (Subowo, 1990).
Lisosom memainkan peranan yang sangat penting dalam resorbsi tulang yang dilakukan oleh osteoklas (gambar 7.10)
Selain itu, lisosom memegang peranan penting di dalam sekresi kelenjar tiroid oleh sel-sel epitel dari folikel tiroid.
kelenjar tiroid (Thorpe, 1984)
Lisosom memainkan peranan yang sangat penting selama berlangsungnya fertilisasi pada berbagai jenis hewan termasuk manusia, terutama selama berlang-sungnya reaksi akrosom. Enzim-enzim yang dilepaskan dari vesikula akrosom melakukan pencernaan terhadap selaput-selaput pelindung telur sehingga memungkinkan sel pronuklei jantan masuk menembus membran telur untuk berfusi dengan pronuklei betina .

PENDAHULUAN
Badan golgi merupakan organel berbatas membran di dalam sel eukariota yang berperan dalam berbagai fungsi penting, meliputi (i) mengemas bahan-bahan sekresi yang dilepaskan dari sel, (ii) pemrosesan protein meliputi glikosilasi, fosforilasi, sulfasi dan proteolisis selektif, (iii) tempat utama sintesis karbohidrat, menyortir dan mendistribusikan produkproduk retikulum endoplasma (Sheeler dan Bianchii, 1983; Allar, 2005), (iv) sintesis glikolipida, dan (v)proliferasi elemen-elemen membran untuk membran plasma (Sheeler dan Bianchii, 1983)
Badan golgi ditemukan pada tahun 1898 oleh C. Golgi. Ia melaporkan adanya struktur-struktur internal yang dijumpai di dalam sel saraf yang olehnya diberi nama Apparato Reticular Interno atau struktur retikular internal (Thorpe, 1984). Atas dasar penemuan tersebut, pada tahun 1906, C, Golgi menerima hadiah nobel dalam bidang sitologi (Sheeler dan Bianchii, 1983). Kompleks golgi memiliki banyak nama, antara lain diktiosom, golgisom, lipokondria, idiosom, badan golgi, zat golgi, aparat golgi, kompleks golgi, dan vesikula golgi.
B. STRUKTUR BADAN GOLGI
Pengamatan dengan mikroskop elektron menunjukkan bahwa badan golgi tampak menyerupai kantung-kantung pipih dengan sejumlah struktur-struktur perifer yang bervariasi. Setiap kantung pipih diberi nama sakula atau lamella atau sisterna. Setiap sakula berbatas membran dengan tebal kurang lebih 7,5 nm dan di dalamnya terdapat ruang dengan lebar berkisar 15 nm yang diberi nama lumen (Sheeler dan Bianchii, 1983).
Pada sel tumbuhan, tumpukan sejumlah lamella dinama-kan diktiosom. Jarak antar lamella dalam suatu diktiosom berkisar 20 nm. Jumlah lamella pada suatu diktiosom kurang lebih 10 buah. Permukaan kompleks golgi yang terorientasi ke arah retikulum endoplasma disebut permukaan pembentukan atau permukaan cis. Sedang-kan permukaan yang lain disebut permukaan matang atau permukaan trans yang terorientasi ke arah membran plasma (Sheeler dan Bianchii, 1983).
Sisterna pada permukaan pembentukan berbentuk cembung, sedangkan sisterna pada permukaan matang berbentuk cekung. Vesikula-vesikula sederhana yang berada disekitar permukaan pembentukan akan berfusi dan berkontribusi menambah struktur badan golgi. Vesikulavesikula yang terdapat di sekitar permukaan matang lebih besar dan dibentuk dari permukaan sisterna. Vesikula-vesikula sederhana juga dilepaskan dari bagian tepi sisterna diantara permukaan pemebentukan dan permukaan matang.
Badan golgi pada kebanyakan sel terutama berfungsi dalam hubungannya dengan fungsi sekresi. Permukaan pembentukan yang terletak di dekat inti atau di dekat bagian khusus dari retikulum endoplasma yang tidak memiliki ribosom dinamakan retikulum endoplasma transisi. Membran inti dan retikulum endoplasma halus adalah sumber vesikula-vesikula sederhana yang berfusi dengan permukaan pembentukan. Beberapa vesikula-vesikula besar dibentuk dari permukaan matang dinamakan vesikula sekresi dan kelak akan berfusi dengan membran plasma. Jika vesikula-vesikula dilepaskan dari permukaan matang badan golgi, juga memungkinkan terbentuknya struktur-struktur internal sel seperti yang terjadi selama pembentukan akrosom pada sel sperma. Atau pembentukan lisosom (Sheler dan Bianchii, 1983.
B. LINTASAN SEKRESI
Secara morfologis, kompleks golgi menunjukkan suatu polaritas yang terkait secara langsung dengan fungsi organel. Di dalam sel-sel sekresi, kompleks golgi merupakan suatu pusat fungsional dalam rantai sekresi. Elemen-elemen penghubung lintasan sekresi tersebut adalah retikulum endoplasma, vesikula transisi, kompleks golgi, vesikula sekresi, dan membran plasma. Lintasan rantai sekresi ditunjukkan pada gambar 8.3.
Lintasan skretori meliputi minimal 6 tahap, yaitu (i) tahap sintesis, (ii) tahap segregasi, (iii) tahap transpor intraseluler, (iv) tahap konsentrasi, (v) tahap penyimpanan, dan (vi) tahap pengeluaran. Selama berlangsungnya sekresi, terjadi aliran materi dari retikulum endopla-sma ke arah badan golgi. Permukaan cekung badan golgi yang terorientasi ke arah retikulum endoplasma disebut permukaan pembentukan atau permukaan cis, sedangkan permukaan cembung yang terorientasi ke arah permukaan sel disebut permukaan matang atau permukaan trans.
Pada permukaan matang badan golgi, dibentuk vesikulavesikula sekresi, sedangkan pada permukaan pembentukan terdapat vesikula-vesikula transpor yang dibentuk dari membrane retikulum endoplasma. Selama berlangsungnya proses sekresi, terjadi aliran materi yang dibentuk dalam retikulum endoplasma dan bergerak melalui permukaan badan golgi dari permukaan pembentukan ke permukaan matang. Selanjutnya bergerak sebagai granula-granula sekresi pada daerah apikal sel dan pada akhirnya menuju ke membran plasma dimana bahanbahan tersebut dikeluarkan.
C. ASAL BADAN GOLGI
Menurut (Sheeler dan Bianchii, 1983) ada tiga sumber yang diusulkan yang diduga sebagai asal badan golgi, yaitu:
1. Vesikula-vesikula yang berasal dari membran luar salut inti atau retikulum endoplasma.
2. Vesikula-vesikula atau struktur-struktur sitoplasma yang lain.
3. Pembelahan dari badan golgi yang telah ada di dalam sel.
Sisterna dari golgi dapat dibentuk dari vesikula-vesikula yang berasal dari membran luar salut inti atau reticulum endoplasma. Vesikula-vesikula transisi bermigrasi ke permukaan pembentukan dari badan golgi dan selanjutnya berfusi dengan membran sisterna badan golgi yang sudah ada. Dengan cara ini, satu kompleks golgi dapat dibentuk secara sempurna.
Agregasi-agregasi dari vesikula transisi terjadi pada daerah sitoplasma yang disebut zona eksklusi (zones of exclusion) yang bebas ribosom. Zona tersebut biasanya dikelilingi oleh membran-membran retikulum endoplasma atau membran inti. Badan golgi-badan golgi sederhana diasumsikan telah ada pada awal perkembangan organel yang dijumpai pada zona tersebut. Beberapa bukti bahwa badan golgi terbentuk pada zona eksklusi (Sheeler dan Bianchii, 1983) adalah sebagai berikut:
1. Sel-sel pada biji yang dorman pada umumnya tidak memiliki badan golgi, namun pada zona eksklusi terdapat kumpulan vesikula-vesikula kecil. Hasil fotomikrograf pada sel biji pada stadium awal perkecambahan mendukung adanya per-kembangan badan golgi yang progresif pada zona eksklusi. Perkembangan badan golgi ber-tepatan dengan hilangnya vesikula-vesikula tran-sisi.
2. Pada telur katak, badan golgi tampak berkembang dari kelompok-kelompok vesikula yang terdapat pada zona eksklusi.
Selama berlangsungnya pembelahan sel pada sel hewan dan tumbuhan, jumlah badan golgi meningkat. Jumlah badan golgi yang dijumpai pada sel anak setelah pembelahan sama dengan jumlah badan golgi pada sel induk. Pada algae Botrydium granulatum yang multinukleat, pada setiap kutub sel yang sedang membelah tepat pada pembentukan spindel, terdapat sebuah badan golgi. Pada metafase lanjut, kedua badan golgi tersebut terdapat pada setiap ujung spindel dan dipisahkan oleh sentriol. Hal tersebut mendukung bahwa badan golgi dapat dibentuk melalui pembelahan organel.
Pembentukan badan golgi dari retikulum endoplasma melalui peleburan sisterna yang berbentuk tubular yang terdapat pada bagian perifer dari retikulum endop-lasma kasar. Hal ini terlihat pada sel-sel hati embrio. Pada saat sel-sel hati matang, sisterna tubular berubah menjadi bentuk mangkuk, kemudian bagian luar dari sisterna dibentuk vesikula-vesikula sekresi. Fenomena yang se-rupa dijumpai pada zoospora dan pembentukan sperma (gambar 7.4).
D. KOMPOSISI KIMIA BADAN GOLGI
Seperti halnya retikulum endoplasma, badan golgi juga mengandung senyawa lipida yang terdiri atas fosfolipida dan lemak netral. Sedangkan protein terdiri atas glikoprotein, mykoprotein, dan enzim.
1. Kandungan Lemak
Membran badan golgi memiliki kandungan lipida yang berbeda dari membran sel lain (tabel 8.1)
2. Kandungan Enzim
Analisa enzimatik dari badan golgi yang diisolasi didapatkan bahwa, badan golgi mengandung enzim-enzim yang sangat heterogen, misalnya enzim-enzim glikolisil transferase, oksidoreduktase, fosforase, dan sebagainya (tabel 8.2).
E. FUNGSI BADAN GOLGI
Seperti diungkapkan sebelumnya bahwa, fungsi dari kompleks golgi adalah sebagai tempat pemrosesan protein pasca translasi. Protein-protein tersebut berasal dari lumen retikulum endoplasma dan bergerak ke badan golgi. Ada dua cara bagaimana protein melewati permukaan pembentukan ke permukaan matang dari badan golgi, yaitu (i) Model Sisternal Progression atau model transport vesikula dan (ii) Model Sisternal Transfer atau model pematangan sisterna (Thorpe, 1984; Allar, 2005)
Pada model sisternal progression, vesikula-vesikula yang berisi protein yang berasal dari retikulum endoplasma berfusi dengan permukaan pembentukan dari badan golgi untuk mengalami proses lebih lanjut. Kemudian setiap sisterna bergerak melalui tumpukan badan golgi ke arah permukaan trans. Pada permukaan trans, sisterna dipecah-pecah menjadi sejumlah vesikula kecil yang membawa protein ke tujuannya. Kelemahan model ini adalah karena sisterna badan golgi mempunyai sifat-sifat yang berbeda, sehingga sulit untuk menjelaskan bagaimana satu sisterna dengan sifat-sifat tertentu dapat berubah menjadi sisterna dengan sifat-sifat lain (Thorpe, 1984; Allar, 2005).
Pada model sisternal transfer, protein bergerak dari satu sisterna ke sisterna yang lain melalui pembentukan vesikulavesikula kecil yang dilepaskan dari sisterna sebelumnya. Vesikula-vesikula tersebut bergerak maju dan berdifusi dengan sisterna berikutnya. (Thorpe, 1984; Allar, 2005)
Kompleks golgi berfungsi dalam biosintesis glikoprotein dan glikolipida. Glikoprotein adalah protein yang mengandung karbohidrat yang terikat secara kovalen, biasanya berupa Dgalaktosa, D-mannosa, L-fucosa, D-glukosamin, N-asetil-Dgalaktosamin, dan asam N-asetil-muramat atau asam sialat. Unit-unit monosakarida tersebut terikat dalam rantai oligosakarida.
Bahan-bahan yang akan disekresikan pada akhirnya berkumpul pada permukaan trans badan golgi dan kemudian dilepaskan dalam bentuk vesikula. Vesikula-vesikula sekresi melepaskan kandungannya dengan dua cara, yaitu secara konstitutif dan secara regulatif. Sejumlah protein-protein terlarut maupun yang terikat membran yang baru disintesis, lipida membran plasma yang baru disintesis dilepaskan dengan cara konstitutif, artinya tidak tergantung pada signal-signal tertentu seperti hormone atau neurotransmitter. Sejumlah proteinprotein tertentu yang tersimpan di dalam vesikula sekresi hanya dapat dilepaskan bilamana ia menerima sinyal-sinyal tertentu yang berasal dari hormone atau neurotransmitter. Sekresi seperti ini dinamakan sekresi regulative.
1. Sintesis, Pengemasan dan Pelepasan Hormon Peptida
Sintesis, pengemasan dan pelepasan hormon-hormon peptida melibatkan organel-organel sitoplasmik, yaitu reticulum endoplasma kasar bersama ribosom dan badan golgi. Tahap awal adalah sintesis protein pada yang berlangsung pada ribosom yang melekat pada retikulum endoplasma dan menghasilkan rantai polipeptida yang dikenal sebagai preprohormon. Rantai polipeptida tersebut diarahkan ke dalam lumen reticulum endoplasma oleh signal sequence asam-asam amino. Enzim-enzim pada retikulum endoplasma memotong signal sequence dan menghasilkan prohormon yang tidak aktif. Prohormon selanjutnya ditranspor menuju badan golgi. Di dalam badan golgi dikemas dan dilepaskan melalui pertunasan badan golgi dalam bentuk vesikula sekresi.
Di dalam vesikula sekresi terdapat sejumlah enzim yang berperan memotong prohormon menjadi satu atau lebih hormon yang aktif dan sejumlah fragemen-fragmen peptida. Hormon-hormon tersebut selanjutnya dirembeskan masuk ke dalam aliran darah untuk dibawah menuju jaringan target.
2. Sekresi Kelenjar Tiroid
Satu peranan kompleks golgi dalam biosintesis glikoprotein terlihat jelas pada sel-sel folikel tiroid. Sel-sel ini terlibat dalam sintesis tiroglobulin, suatu glikoprotein yang disekresikan oleh sel-sel ke dalam folikel dimana ia diiodinasi dan disimpan. Folikel adalah ruang interseluler yang besar dan dikelilingi oleh sel-sel epitel yang mensintesis tiroglobulin.
Tiroglobulin terdiri atas dua tipe rantai polipeptida yaitu tipe pertama yang hanya terdiri atas disakarida Nasetilglukosamin yang terikat pada mannosa dan tipe kedua yang lebih kompleks. Selain mengandung gula, juga mengandung galaktosa, fruktosa, dan asam sialat (Thorpe, 1984)
Urutan selama pembentukan tiroglobulin ditunjukkan pada gambar 8.6. Tiroglobulin diangkut secara vektorial dalam sel, dimulai pada retikulum endoplasma kasar dimana rantai polipeptida dibentuk. Polipetida mengalami prosesing berupa penambahan unit-unit monosakarida membentuk glikoprotein yang belum sempurna. Glikoprotein yang belum sempurna dibawa ke badan golgi melalui vesikula-vesikula transpor. Di dalam badan golgi glikoprotein mengalami penyempurnaan dengan penambahan unit-unit galaktosa (Thorpe, 1984).
Vesikula-vesikula sekresi yang mengandung glikoprotein yang telah sempurna dibentuk secara apikal dari kompleks golgi dan bergerak ke arah lumen folikel. Di dalam folikel berlangsung iodinasi. Glikoprotein yang telah diiodinasi kemudian disimpan dalam folikel-folikel hingga sel epitel distimulasi oleh TSH dari kelenjar pituitari untuk mengabsorbsi tiroglobulin. Hormon tiroid yang telah diiodinasi dipotong-potong dan selanjutnya dilepaskan ke dalam darah untuk selanjutnya menuju organ target.
Sel-sel folikel tiroid secara aktif mentranspor ion-ion iodine dari darah. Ion-ion tersebut selanjunya ditranspor menuju lumen folikel tiroid. Beberapa asam amino tirosin di dalam tiroglobulin akan mengalami iodinasi. Namun demikian ion-ion iodin tidak dapat terikat secara langsung pada asam amino tirosin. Oleh sebab itu ion-ion iodin harus melepaskan elektronnya melalui proses oksidasi. Oksidasi ion-ion iodin dikatalisis oleh enzimenzim peroksidase. 1 atau 2 ion-ion iodin yang telah mengalami oksidasi selanjutnya berikatan dengan asam amino tirosin membentuk T1 dan T2. T1 selanjuntnya bergandengan dengan T2 membentu T3 dan T2 bergandengan dengan T2 membentu T4.
Tiroglobulin yang telah mengalami iodinasi tetap berada di dalam folikel tiroid dalam bentuk koloid. Dibawah pengaruh hormone Thyroid stimulating hormon (TSH) yang berasal dari hipofisis, tiroglobulin selanjutnya dimasukkan di dalam sel-sel folikel tiroid secara endositosis. Di dalam sel-sel folikel, endosom yang mengandung tiroglobulin berfusi dengan lisosom primer dan selanjutnya berlangsung proses pemotongan. Pemotongan tiroglobulin tersebut oleh enzim-enzim lisosom menghasilkan hormone triiodotironin (T3) dan tetraiodotironin (T4). Selanjutnya hormone tersebut siap dirembeskan ke aliran darah.
3. Proliferasi membran-membran seluler
Kompleks golgi selain berperan dalam sekresi, juga memainkan peranan dalam persiapan protein-protein untuk organel-organel seperti lisosom dan membran plasma (gambar 8.8). Protein -protein yang dipersiapkan untuk lisosom atau membran plasma disintesis oleh ribosom-ribosom yang melekat pada retikulum endoplasma kasar. Beberapa dari proteinprotein tersebut dilepaskan ke dalam lumen reticulum endoplasma dan yang lain tetap pada membran reticulum endoplasma dan kelak menjadi dinding vesikula transpor. Dalam beberapa menit setelah sintesis, protein-protein tersebut tampak pada permukaan cis dari badan golgi.
Mekanise transport protein dari lumen RE ke badan golgi berlangsung melalui vesikula transport. Vesikula transport berfusi dengan permukaan cis badan golgi. Selanjutnya protein berpindah dari suatu sisterna ke sisterna berikutnya hingga mencapai permukaan trans. Protein-protein yang dipersiapkan untuk menjadi komponen membran lisosom dan membrane plasma tetap tertanam pada membrane RE. Protein tersebut diranspor dengan cara yang sama, namun pada saat tiba pada sasaran, protein tersebut tetap terikat pada membran.
4. Menyortir Protein-protein untuk Sasaran Tertentu
Protein-protein yang disintesis pada reticulum endoplasma yang diperuntukkan untuk membran plasma, lisosom, dan vesikula sekresi, diangkut ke badan golgi pada permukaan pembentukan. Protein-protein yang sampai pada permukaan pembentukan kompleks golgi bersama-sama dengan protein membran retikulum endoplasma. Vesikulavesikula yang dilepaskan dari permukaan trans tidak mengandung protein membran retikulum endoplasma. Kompleks golgi berperan memilih protein membran reticulum endoplasma oleh vesikula-vesikula kecil dari sisterna badan golgi permukaan pembentukan.
Rothman (1981 dalam Thorpe, 1984) membedakan permukaan pembentukan badan golgi dengan permukaan matang badan golgi. Permukaan pembentukan terdiri atas semua sisterna golgi kecuali satu atau dua yang terakhir. Peranan permukaan pembentukan adalah memilih protein retikulum endoplasma yang akan dikembalikan ke reticulum endoplasma. Permukaan matang dari badan golgi terdiri atas ½ sisterna permukaan yang berperan menerima protein yang telah dimurnikan dan menyebarkan melalui vesikula-vesikula ke lokasinya yang tepat di dalam sel. Rothman (1981) mengusulkan bahwa badan golgi terdiri atas tiga kompartemen yaitu kompartemen cis (kompartemen pembentukan), kompartemen medial, dan kompartemen trans (kompartemen matang). Kompartemen cis memilih dan melepaskan protein-protein retikulum endoplasma dan juga menambah gugus fosfat ke gula terminal protein lisosom. Kompartemen medial (terdiri atas sisterna di tengah-tengah tumpukan golgi) merupa-kan tempat penambahan Nasetilglukosamin. Sedangkan kompartemen trans merupakan tempat penambahan unit-unit galaktosa dan asam sialat, juga memilih berbagai protein sesuai dengan tujuan akhirnya. Penambahan gugus fosfat pada gula terminal protein lisosom dalam kompleks tersebut untuk mencegah penambahan Nasetilglukosamin dalam kompartemen medial dan penambahan galaktosa dan asam sialat dalam kompartemen trans.
5. Pembentukan Dinding Sel
Badan golgi berperan dalam pembentukan papan sel dan dinding sel. Papan sel dan dinding sel terbentuk selama anafase dan telofase mitosis dan miosis kedua. Sebelum anafase, kompleks golgi berada di luar kumparan. Selama anafase, kompleks golgi melepaskan vesikula-vesikula menuju pusat kumparan dan menimbun disekitar benang-benang kumparan. Vesikula berisi senyawa pembentuk papan sel dan dinding sel.
Vesikula-vesikula yang berasal dari badan golgi berisi bahan untuk pembentukan papan sel dan dinding sel. Di dalam sitoplasma terdapat mikrotubul-mikrotubul yang tersusun parallel dan disebut fragmoplas. Vesikula-vesikula yang ber asal dari badan golgi berhubungan dengan fragmoplas dan ditransportasikan sepanjang mikrotubul ke arah ekuatorial dan terakumulasi pada daerah dimana mikrotubul mengalami overlapping. Vesikula-vesikula yang mengandung bahan untuk papan sel dan dinding sel diakumulasikan pada daerah ekuatorial pada mikrotubul yang tumpang tindih dan berfusi membentuk papan sel. Bahan-bahan dari vesikula bergabung membentuk dinding sel. Vesikula-vesikula golgi yang baru terbentuk diakumulasi pada bagian tepi papan sel, kemudian berfusi dan meluas ke arah luar. Membran papan sel yang sedang merentang berfusi dengan membran plasma. Bahanbahan dinding sel dideposisikan membentuk dinding sel yang sempurna (Albert et al., 1983).
6. Pembentukan Akrosom
Akrosom adalah suatu badan berbatas membran yang terletak pada bagian kepala spermatozoa di sebelah anterior dari inti. Badan golgi merupakan organel yang berperan dalam pembentukan akrosom melalui beberapa fusi vesikula-vesikula yang mengandung bahan untuk akrosom. Sedangkan membrane vesikula-vesikula menjadi membran akrosom. Akrosom mengandung berbagai jenis enzim-enzim hidrolitik yang penting di dalam proses fertilisasi. Tanpa akrosom, maka sperma mengalami kesulitan untuk menembus berbagai selaput yang melindungi sel telur. Proses pembentukan akrosom pada kepala sperma.
STRUKTUR SEL TUMBUHAN DAN HEWAN & MEKANISME TRANSPOR ZAT MELALUI MEMBRAN-
Pada materi ini kita akan mempelajari bagaimana struktur sel tumbuhan?
bagaimana struktur sel hewan ? apa perbedaan sel tumbuhan dan sel hewan?
dan bagaimana mekanisme transpor zat melalui membran sel?
STRUKTUR SEL TUMBUHAN DAN HEWAN
dapat terlihat bahwa pada bagian luar sel tumbuhan terdapat dinding
sel, tetapi pada hewan tidak. Adanya dinding sel inilah yang
menyebabkan selsel tumbuhan memiliki sifat keras dan kaku. Pada
tumbuhan, dinding sel berfungsi antara lain untuk melindungi protoplas,
sebagai penguat tanaman dan mencegah terjadinya dehidrasi. Komponen
utama penyusun dinding sel adalah polisakarida. Dinding sel tumbuhan
muda masih terlihat tipis yang terdiri atas selaput zat pektin. Setelah
sel tumbuhan bertambah tua, maka dinding sel akan menebal dan zat
pembentuknya adalah selulosa. Dinding sel bagian dalam berhubungan
langsung dengan membran plasma. Membran ini bisa terlihat apabila sel
berada di dalam larutan yang lebih pekat daripada larutan dalam sel,
sehingga membran plasma akan lepas.

Selain itu, organela khas yang terdapat pada sel hewan dan tidak
banyak terdapat pada sel tumbuhan adalah sentrosom dan lisosom. Struktur
sentrosom bisa dilihat dengan jelas apabila menggunakan mikroskop
elektron. Struktur sentrosom seperti rakitan batang-batang yang menyusun
bangunan sekunder, sembilan batang membentuk bangun silinder, satu
batang berada di pusat silinder. Apakah dan bagaimanakah lisosom itu?
Cobalah ingat kembali pengertiannya berdasarkan penjelasan di depan.
Secara garis besar, perbedaan antara struktur hewan dengan tumbuhan
bisa dilihat pada Tabel 1.1 di bawah ini! 
Berdasarkan penjelasan tersebut kita dapat membedakan kondisi antara sel hewan dan tumbuhan
MEKANISME TRANSPOR ZAT MELALUI MEMBRAN
Dari penjelasan di depan Anda telah mengetahui bahwa sel merupakan
penyusun jaringan tumbuhan dan hewan. Segala aktivitas terjadi
dalam sel, sehingga fungsi jaringan pun dapat dilakukan dengan baik.
Tentunya di sini ada hubungan antara sel satu dengan yang lain, terutama
dalam hal transpor zat-zat untuk proses metabolisme tumbuhan. Zat-zat
tersebut keluar masuk sel dengan melewati membran sel. Cara zat melewati
membran sel melalui beberapa mekanisme berikut.
1. Transpor Pasif
Transpor pasif merupakan perpindahan zat yang tidak memerlukan
energi. Perpindahan zat ini terjadi karena perbedaan konsentrasi
antara zat atau larutan. Transpor pasif melalui peristiwa difusi,
osmosis, dan difusi terbantu.
a. Difusi
Proses ini merupakan perpindahan molekul larutan berkonsentrasi
tinggi menuju larutan berkonsentrasi rendah tanpa melalui selaput
membran.

menunjukkan proses terjadinya difusi. Pada permulaan percobaan semula molekul glukosa ada di bagian A.
Setelah beberapa saat, proses difusi menyebabkan konsentrasi
glukosa di A turun dan di B naik dengan kecepatan yang sama. Setelah 3
jam, konsentrasi pada kedua ruang tersebut sama dan keseimbangan akan
tercapai. Difusi pada membran sel (selaput plasma) dapat terlihat pada

Proses difusi sering terjadi pada tubuh kita. Tanpa kita sadari,
tubuh kita selalu melakukan proses ini, yaitu pada saat kita menghirup
udara. Ketika menghirup udara, di dalam tubuh akan terjadi pertukaran
gas antarsel melalui proses difusi. Contoh lain proses difusi adalah
saat kita membuat minuman sirup. Sirup yang kita larutkan dengan air
akan bergerak dari larutan yang konsentrasinya tinggi ke larutan
yang konsentrasinya rendah. Pada masing-masing zat, kecepatan difusi
berbeda-beda. Untuk contoh kasus yang dijelaskan, yaitu antara sirup dan
gas, maka kecepatan difusi sirup lebih besar pada gas. Berdasarkan hal
itu coba Anda cari lagi contoh proses difusi lain yang pernah Anda
ketahui!
b. Osmosis
Untuk memahami tentang osmosis, perhatikan Gambar 1.16! Gambar 1.16
menunjukkan proses osmosis. Air akan berpindah dari A menuju B melalui
membran semi permeabel sehingga diperoleh hasil larutan isotonis, yaitu
konsentrasi air sama untuk dua larutan antara A dan B, walaupun hasil
akhirnya nanti volume antara A dan B berbeda. Setelah terjadi osmosis,
maka gambar prosesnya menjadi seperti berikut.

Dari ilustrasi itu dapat disimpulkan bahwa osmosis adalah
proses perpindahan air dari zat yang berkonsentrasi rendah (hipotonis)
ke larutan yang berkonsentrasi tinggi (hipertonis) melalui membran
semipermeabel, sehingga didapatkan larutan yang berkonsentrasi seimbang
(isotonis). Peristiwa osmosis dapat kita temukan dalam kehidupan
sehari-hari antara lain pada penyerapan air melalui bulu-bulu akar, dan
mengerutnya sel darah merah yang dimasukkan ke dalam larutan hipertonis.
Difusi Terbantu
Proses difusi terbantu difasilitasi oleh suatu protein. Difusi
terbantu sangat tergantung pada suatu mekanisme transpor dari membran
sel. Difusi terbantu dapat ditemui pada kehidupan sehari-hari, misalnya
pada bakteri Escherichia coli yang diletakkan pada media laktosa.
Membran sel bakteri tersebut bersifat impermeabel sehingga tidak dapat
dilalui oleh laktosa. Setelah beberapa menit kemudian bakteri akan
membentuk enzim dari dalam sel yang disebut permease, yang merupakan
suatu protein sel. Enzim permease inilah yang akan membuatkan jalan bagi
laktosa sehingga laktosa ini dapat masuk melalui membran sel.
Transpor Aktif
Transpor aktif merupakan transpor partikel-partikel melalui
membran semipermeabel yang bergerak melawan gradien konsentrasi
yang memerlukan energi dalam bentuk ATP. Transpor aktif berjalan dari
larutan yang memiliki konsentrasi rendah ke larutan yang memiliki
konsentrasi tinggi, sehingga dapat tercapai keseimbangan di dalam
sel. Adanya muatan listrik di dalam dan luar sel dapat mempengaruhi
proses ini, misalnya ion K+, Na+dan Cl+. Peristiwa transpor aktif dapat
Anda lihat pada peristiwa masuknya glukosa ke dalam sel melewati membran
plasma dengan menggunakan energi yang berasal dari ATP. Contoh lain
terjadi pada darah di dalam tubuh kita, yaitu pengangkutan ion kalium
(K) dan natrium (Na) yang terjadi antara sel darah merah dan cairan
ekstrasel (plasma darah). Kadar ion kalium pada sitoplasma sel
darah merah tiga puluh kali lebih besar daripada cairan plasma darah.
Tetapi kadar ion natrium plasma darah sebelas kali lebih besar daripada
di dalam sel darah merah. Adanya pengangkutan ion bertujuan agar dapat
tercapai keseimbangan kadar ion di dalam sel. Mekanisme transpor ion

Peristiwa transpor aktif dibedakan menjadi dua, yaitu endositosis dan eksositosis.
Endositosis
Endositosis merupakan peristiwa pembentukan kantong membran
sel. Endositosis terjadi karena ada transfer larutan atau partikel ke
dalam sel. Peristiwa endositosis dibedakan menjadi dua, yaitu sebagai
berikut.
Pinositosis
Pinositosis merupakan peristiwa masuknya sejumlah kecil medium
kultur dengan membentuk lekukan-lekukan membran sel. Peristiwa ini dapat
terjadi bila konsentrasi protein dan ion tertentu pada medium
sekeliling sel sesuai dengan konsentrasi di dalam sel. Proses
pinositosis dapat diamati dengan mikroskop elektron. Sel-sel yang
melakukan proses pinositosis ini antara lain sel darah putih, epitel
usus, makrofag hati, dan lain-lain. Tahapan proses pinotosis
adalah sebagai berikut.

Keterangan gambar:
1. Molekul-molekul medium kultur mendekati membran sitoplasma.
2. Molekul-molekul mulai melekat (menempel) pada plasma, hal ini
terjadi karena adanya konsentrasi yang sesuai antara protein dan ion
tertentu pada medium sekeliling sel dengan di dalam sel.
3. Mulai terbentuk invaginasi pada membran sitoplasma.
4. Invaginasi semakin ke dalam sitoplasma.
5. Terbentuk kantong dalam sitoplasma dan saluran pinositik.
6. Kantong mulai lepas dari membran plasma dan membentuk gelembunggelembung kantong.
7. Gelembung-gelembung kantong mulai mempersiapkan diri untuk melakukan fragmentasi.
8. Gelembung pecah menjadi gelembung yang lebih kecil.
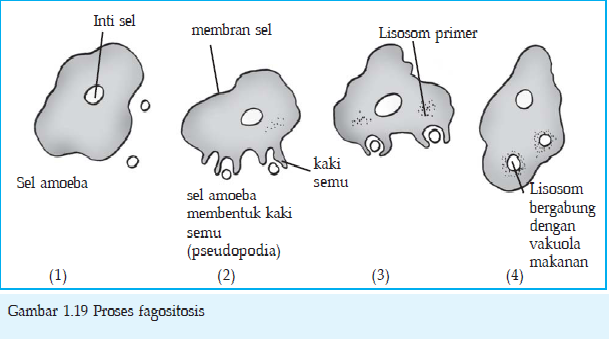
Fagositosis
Fagositosis merupakan peristiwa yang sama seperti pada pinositosis
tetapi terjadi pada benda padat yang ukurannya lebih besar. Fagositosis
dapat diamati dengan mikroskop misalnya yang terjadi pada
Amoeba.Keterangan gambar:
1. Sebuah sel Amoeba mendekati sel Paramaecium.
2. Amoeba membentuk kaki semu (pseudopodia) dan semakin mendekati Paramaecium.
3. Amoeba mengurung sel Paramaecium dengan kaki semu dan memasukkannya ke dalam vakuola makanan.
4. Lisosom pada Amoeba mulai bergabung (fusi) dengan vakuola makanan untuk mengeluarkan enzim pencernaan.
Eksositosis
Eksositosis adalah proses keluarnya suatu zat ke luar sel. Proses
ini dapat Anda lihat pada proses kimia yang terjadi dalam tubuh kita,
misalnya proses pengeluaran hormon tertentu. Semua proses sekresi dalam
tubuh merupakan proses eksositosis. Sel-sel yang mengeluarkan protein
akan berkumpul di dalam badan golgi. Kantong yang berisi protein akan
bergerak ke arah permukaan sel untuk mengosongkan isinya.
Tahap-tahap fagositosis
Mitokondria (mitochondrion’, plural: mitochondria’) atau kondriosom (chondriosome) adalah organela tempat berlangsungnya fungsi responsi sel makhluk hidup. Respirasi merupakan proses perombakan atau katabolisme untuk menghasilkan energi atau tenaga bagi berlangsungnya proses hidup. Dengan demikian, mitokondria adalah “pembangkit tenaga” bagi sel.
Mitokondria merupakan salah satu bagian sel yang paling penting karena di sinilah energi dalam bentuk ATP [Adenosine Tri-Phosphate] dihasilkan. Mitokondria mempunyai dua lapisan membran, yaitu lapisan membran luar dan lapisan membran dalam. Lapisan membran dalam ada dalam bentuk lipatan-lipatan yang sering disebut dengan cristae. Di dalam Mitokondria terdapat ‘ruangan’ yang disebut matriks, dimana beberapa mineral dapat ditemukan. Sel yang mempunyai banyak Mitokondria dapat dijumpai di jantung, hati, dan otot.
Keberadaan mitokondria didukung oleh hipotesis endopsimbiosis yang mengatakan bahwa pada tahap awal evolusi sel eukariot bersimbiosis dengan prokariot (bakteri) [Margullis, 1981]. Kemudian keduanya mengembangkan hubungan simbiosis dan membentuk organel sel yang pertama. Adanya DNA pada mitokondria menunjukkan bahwa dahulu mitokondria merupakan entitas yang terpisah dari sel inangnya. Hipotesis ini ditunjang oleh beberapa kemiripan antara mitokondria dan bakteri. Ukuran mitokondria menyerupai ukuran bakteri, dan keduanya bereproduksi dengan cara membelah diri menjadi dua. Hal yang utama adalah keduanya memiliki DNA berbentuk lingkar. Oleh karena itu, mitokondria memiliki sistem genetik sendiri yang berbeda dengan sistem genetik inti. Selain itu, ribosom dan rRNA mitokondria lebih mirip dengan yang dimiliki bakteri dibandingkan dengan yang dikode oleh inti sel eukariot [Cooper, 2000].
Secara garis besar, tahap respirasi pada tumbuhan dan hewan melewati jalur yang sama, yang dikenal sebagai daur atau Siklus Krebs

 Struktur umum suatu mitokondrion
Struktur umum suatu mitokondrion
Mitokondria banyak terdapat pada sel yang memilki aktivitas
metabolisme tinggi dan memerlukan banyak ATP dalam jumlah banyak,
misalnya sel otot jantung. Jumlah dan bentuk mitokondria bisa
berbeda-beda untuk setiap sel. Mitokondria berbentuk elips dengan
diameter 0,5 µm dan panjang 0,5 – 1,0 µm. Struktur mitokondria terdiri
dari empat bagian utama, yaitu membran luar, membran dalam, ruang antar
membran, dan matriks yang terletak di bagian dalam membran [Cooper,
2000].
Membran luar terdiri dari protein dan lipid dengan perbandingan yang sama serta mengandung protein porin yang menyebabkan membran ini bersifat permeabel terhadap molekul-molekul kecil yang berukuran 6000 Dalton. Dalam hal ini, membran luar mitokondria menyerupai membran luar bakteri gram-negatif. Selain itu, membran luar juga mengandung enzim yang terlibat dalam biosintesis lipid dan enzim yang berperan dalam proses transpor lipid ke matriks untuk menjalani β-oksidasi menghasilkan Asetil KoA.
Membran dalam yang kurang permeabel dibandingkan membran luar terdiri dari 20% lipid dan 80% protein. Membran ini merupakan tempat utama pembentukan ATP. Luas permukaan ini meningkat sangat tinggi diakibatkan banyaknya lipatan yang menonjol ke dalam matriks, disebut krista [Lodish, 2001]. Stuktur krista ini meningkatkan luas permukaan membran dalam sehingga meningkatkan kemampuannya dalam memproduksi ATP. Membran dalam mengandung protein yang terlibat dalam reaksi fosforilasi oksidatif, ATP sintase yang berfungsi membentuk ATP pada matriks mitokondria, serta protein transpor yang mengatur keluar masuknya metabolit dari matriks melewati membran dalam.
Ruang antar membran yang terletak diantara membran luar dan membran dalam merupakan tempat berlangsungnya reaksi-reaksi yang penting bagi sel, seperti siklus Krebs, reaksi oksidasi asam amino, dan reaksi β-oksidasi asam lemak. Di dalam matriks mitokondria juga terdapat materi genetik, yang dikenal dengan DNA mitkondria (mtDNA), ribosom, ATP, ADP, fosfat inorganik serta ion-ion seperti magnesium, kalsium dan kalium
Mitokondria merupakan salah satu bagian sel yang paling penting karena di sinilah energi dalam bentuk ATP [Adenosine Tri-Phosphate] dihasilkan. Mitokondria mempunyai dua lapisan membran, yaitu lapisan membran luar dan lapisan membran dalam. Lapisan membran dalam ada dalam bentuk lipatan-lipatan yang sering disebut dengan cristae. Di dalam Mitokondria terdapat ‘ruangan’ yang disebut matriks, dimana beberapa mineral dapat ditemukan. Sel yang mempunyai banyak Mitokondria dapat dijumpai di jantung, hati, dan otot.
Keberadaan mitokondria didukung oleh hipotesis endopsimbiosis yang mengatakan bahwa pada tahap awal evolusi sel eukariot bersimbiosis dengan prokariot (bakteri) [Margullis, 1981]. Kemudian keduanya mengembangkan hubungan simbiosis dan membentuk organel sel yang pertama. Adanya DNA pada mitokondria menunjukkan bahwa dahulu mitokondria merupakan entitas yang terpisah dari sel inangnya. Hipotesis ini ditunjang oleh beberapa kemiripan antara mitokondria dan bakteri. Ukuran mitokondria menyerupai ukuran bakteri, dan keduanya bereproduksi dengan cara membelah diri menjadi dua. Hal yang utama adalah keduanya memiliki DNA berbentuk lingkar. Oleh karena itu, mitokondria memiliki sistem genetik sendiri yang berbeda dengan sistem genetik inti. Selain itu, ribosom dan rRNA mitokondria lebih mirip dengan yang dimiliki bakteri dibandingkan dengan yang dikode oleh inti sel eukariot [Cooper, 2000].
Secara garis besar, tahap respirasi pada tumbuhan dan hewan melewati jalur yang sama, yang dikenal sebagai daur atau Siklus Krebs
Struktur

Membran luar terdiri dari protein dan lipid dengan perbandingan yang sama serta mengandung protein porin yang menyebabkan membran ini bersifat permeabel terhadap molekul-molekul kecil yang berukuran 6000 Dalton. Dalam hal ini, membran luar mitokondria menyerupai membran luar bakteri gram-negatif. Selain itu, membran luar juga mengandung enzim yang terlibat dalam biosintesis lipid dan enzim yang berperan dalam proses transpor lipid ke matriks untuk menjalani β-oksidasi menghasilkan Asetil KoA.
Membran dalam yang kurang permeabel dibandingkan membran luar terdiri dari 20% lipid dan 80% protein. Membran ini merupakan tempat utama pembentukan ATP. Luas permukaan ini meningkat sangat tinggi diakibatkan banyaknya lipatan yang menonjol ke dalam matriks, disebut krista [Lodish, 2001]. Stuktur krista ini meningkatkan luas permukaan membran dalam sehingga meningkatkan kemampuannya dalam memproduksi ATP. Membran dalam mengandung protein yang terlibat dalam reaksi fosforilasi oksidatif, ATP sintase yang berfungsi membentuk ATP pada matriks mitokondria, serta protein transpor yang mengatur keluar masuknya metabolit dari matriks melewati membran dalam.
Ruang antar membran yang terletak diantara membran luar dan membran dalam merupakan tempat berlangsungnya reaksi-reaksi yang penting bagi sel, seperti siklus Krebs, reaksi oksidasi asam amino, dan reaksi β-oksidasi asam lemak. Di dalam matriks mitokondria juga terdapat materi genetik, yang dikenal dengan DNA mitkondria (mtDNA), ribosom, ATP, ADP, fosfat inorganik serta ion-ion seperti magnesium, kalsium dan kalium
Fungsi mitokondria
Peran utama mitokondria adalah sebagai pabrik energi sel yang menghasilkan energi dalam bentuk ATP. Metabolisme karbohidrat akan berakhir di mitokondria ketika piruvat di transpor dan dioksidasi oleh O2¬ menjadi CO2 dan air. Energi yang dihasilkan sangat efisien yaitu sekitar tiga puluh molekul ATP yang diproduksi untuk setiap molekul glukosa yang dioksidasi, sedangkan dalam proses glikolisis hanya dihasilkan dua molekul ATP. Proses pembentukan energi atau dikenal sebagai fosforilasi oksidatif terdiri atas lima tahapan reaksi enzimatis yang melibatkan kompleks enzim yang terdapat pada membran bagian dalam mitokondria. Proses pembentukan ATP melibatkan proses transpor elektron dengan bantuan empat kompleks enzim, yang terdiri dari kompleks I (NADH dehidrogenase), kompleks II (suksinat dehidrogenase), kompleks III (koenzim Q – sitokrom C reduktase), kompleks IV (sitokrom oksidase), dan juga dengan bantuan FoF1 ATP Sintase dan Adenine Nucleotide Translocator (ANT) [Wallace, 1997].Siklus Hidup Mitokondria
Mitokondria dapat melakukan replikasi secara mandiri (self replicating) seperti sel bakteri. Replikasi terjadi apabila mitokondria ini menjadi terlalu besar sehingga melakukan pemecahan (fission). Pada awalnya sebelum mitokondria bereplikasi, terlebih dahulu dilakukan replikasi DNA mitokondria. Proses ini dimulai dari pembelahan pada bagian dalam yang kemudian diikuti pembelahan pada bagian luar. Proses ini melibatkan pengkerutan bagian dalam dan kemudian bagian luar membran seperti ada yang menjepit mitokondria. Kemudian akan terjadi pemisahan dua bagian mitokondria [Childs, 1998].DNA mitokondria
Mitokondria memiliki DNA tersendiri, yang dikenal sebagai mtDNA (Ing. mitochondrial DNA). MtDNA berpilin ganda, sirkular, dan tidak terlindungi membran (prokariotik). Karena memiliki ciri seperti DNA bakteri, berkembang teori yang cukup luas dianut, yang menyatakan bahwa mitokondria dulunya merupakan makhluk hidup independen yang kemudian bersimbiosis dengan organisme eukariotik. Teori ini dikenal dengan teori endosimbion. Pada makhluk tingkat tinggi, DNA mitokondria yang diturunkan kepada anaknya hanya berasal dari betinanya saja (mitokondria sel telur). Mitokondria jantan tidak ikut masuk ke dalam sel telur karena letaknya yang berada di ekor sperma. Ekor sperma tidak ikut masuk ke dalam sel telur sehingga DNA mitokondria jantan tidak diturunkan.
Lisosom

Lisosom ditemukan oleh de Duve dkk., pada awal tahun 1950 setelah mempelajari distribusi beberapa jenis enzim yang terlibat di dalam metabolisme karbohidrat. Salah satu enzim yang terlibat di dalam metabolisme karbohidrat. Salah satu enzim yang dipelajari adalah fosfatase asam yang memecah gugus fosfat pada beberapa fosfat yang mengandung ester fosfat.
kelompok lisosom dekat mitokondria
(Sheeler dan Bianchii, 1983)
Lisosom adalah vesikula yang berbatas membran dimana di dalamnya terkandung enzim-enzim hidrolase. Suatu organel dapat didefinisikan sebagai lisosom bilamana memenuhi beberapa kriteria, yaitu:
1. Organel yang bersangkutan berbatas membran;
2. Mengandung dua atau lebih enzim-enzim hidrolase yang semuanya adalah asam hidrolase;
3. Memiliki sifat kelatenan enzim.
Lisosom dibatasi oleh membran tunggal dan di dalamnya terkandung kurang lebih 40 jenis enzim yang semuanya adalah enzim hidrolase seperti protease, nuklease, glikosidase, lipase, fosfolipase, fosfatase, dan sulfatase. Enzim-enzim hidrolase bekerja dengan baik pada pH kurang lebih 5. Untuk mempertahankan pH tersebut, maka secara terus menerus terjadi pemompaan ion hidrogen ke dalam lumen lisosom dengan melibatkan hidrolisis ATP sebagai sumber energi. Lisosom memiliki protein maker yang disebut sebagai “Docking-marker acceptor”. Dengan demikian, lisosom akan dapat berfusi dengan vesikula-vesikula target dengan tepat.
Beberapa molekul sederhana dapat menembus membran lisosom, misalnya quinakrin. Quinakrin dapat meningkatkan pH di dalam lisosom jika diberikan ke dalam sel. Quinakrin digunakan di laboratorium sebagai inhibitor fungsi lisosom. Lisosom memiliki fungi pencernaan intra sel yang sangat luas meliputi pencernaan bahan-bahan intra dan ekstra sel, mikroorganisme yang telah difagositosis dan program kematian sel selama organogenesis oleh sel fagosit (Thorpe, 1984)
Enzim-enzim hidrolase memiliki kemampuan untuk memecahkan berbagai jenis molekul organik seperti polisakarida, protein, lipida, fosfolipida, dan asam-asam nukleat.
B. Pembentukan dan Nasib Lisosom
Asal dan pembentukan lisosom telah dipelajari dengan sangat intensif. Dari berbagai hasil temuan, ada dua pendapat yang berkenaan dengan asal dan pembentukan lisosom, yaitu:
1. Berbagai bukti telah ditemukan bahwa protein-protein hidrolitik dibentuk oleh ribsom yang terdapat pada retikulum endoplasma. Dari retikulum endoplasma kasar, selanjutnya protein tersebut ditranslokasikan menuju permukaan pembentukan badan golgi untuk diproses lebih lanjut. Setelah itu, protein-protein hidrolitik dikemas dan dibungkus dalam bentuk vesikula-vesikula untuk selanjutnya dilepaskan sebagai lisosom primer.
2. Protein-protein hidrolitik dibentuk pada ribosom yang terdapat pada retikulum endoplasma kasar, selanjutnya ia dilepaskan dalam bentuk vesikula menuju daerah GERL (Golgi associated Endoplasmic Reticulum giving rise to Lisosom) yang berdekatan dengan daerah permukaan matang badan golgi. Dari GERL, selanjutnya dilepaskan vesiula-vesikula yang disebut lisosom primer .
Gambar 7.4 Dua ide tentang pembentukan
lisosom (Thorpe, 1984)
Lisosom yang pertama dibentuk oleh sel dan belum terlibat dalam aktivitas pencernaan sel disebut lisosom primer. Sedangkan lisosom sekunder adalah lisosom yang merupakan hasil fusi berulang antara lisosom primer dengan berbagai substrat yang berbatas membran (Albert et al., 1983). Dengan demikian, lisosom sekunder telah terlibat dalam aktivitas pencernaan sel dan di dalam lumennya terdapat substrat dan enzim-enzim hidrolitik. Lisosom sekunder memiliki dua fungsi yang berbeda, yaitu:
1. Heterolisosom, yaitu bila substrat yang dicerna berasal dari luar sel. Dengan demikian, heterolisosom dibentuk dari hasil fusi antara lisosom primer dengan fagosom atau endosom. Heterolisosom sering disebut sebagai vakuola pencerna.
Albert et al. (1983) membagi heterolisosom menjadi dua tipe, yaitu:
- Vakuola pencerna, yaitu hasil fusi antara fagosom (partikelpartikel yang difagositosis seperti bakteri) dengan lisosom primer.
- Badan-badan multivesikula, yaitu hasil fusi antara beberapa endosom (substrat yang masuk secara endositosis dan bukan dalam bentuk partikel) dengan lisosom primer. Dengan demikian, badan-badan multivesikula merupakan kantung-kantung berbatas membran dimana di dalamnya mengandung banyak vesikulavesikula kecil dengan diameter berkisar 50 nm.
Di dalam sel, sesungguhnya terdapat kerjasama yang erat antara heterolisosom dengan autolisosom. Selama heterofagi berlangsung, proteinprotein ditempatkan di dalam vesikula-vesikula endosom, kemudian berfusi dengan lisosom primer dan selanjutnya mengalami hidrolisis. Selama autofagi, sitosegresom berfusi dengan lisosom primer membentuk autolisosom dan memasuki siklus pencernaan intrasel (gambar 7.6)
Tergantung pada keadaan fisiologisnya, vakuola pencerna atau vakuola autofagi pada akhirnya mengalami satu dari tiga kemungkinan yang terjadi, yaitu :
1. Mengosongkan kandungannya dengan cara eksositosis atau defekasi seluler;
2. Menjadi bahan residu tanpa bahan hidrolase;
3. Menghidrolisis kandungannya secara sempurna untuk dapat berdifusi dan selanjutnya siap untuk siklus aktivitas yang baru.
Gambar 7.7. Nasib lisosom (Thorpe, 1984)
Tentang pembentukan sitosegresom, ada beberapa pandangan yang diusulkan, yaitu :
1. Sitosegresom dibentuk dari suatu membran sisterna yang melingkupi mitokondria secara sempurna dan selanjutnya diikuti dengan berdegenerasinya membran dalam;
2. Sitosegresom dibentuk dari suatu membran yang melingkupi mitokondria secara sempurna
3. Sitosegresom dibentuk dari vesikula endosom yang melingkupi mitokondria secara sempurna yang selanjutnya diikuti dengan berdegenerasinya membran dalam;
4. Sitosegresom dibentuk dari vesikula endosom, dimana mitokondria memasuki vesikula endosom melalui suatu celah.
Bentuk akhir heteroslisosom dan autolisoson disebut telolisosom atau postlisosom atau badan residu. Bahan-bahan yang terkandung di dalam telolisosom sewaktu-waktu dapat dilepaskan. Proses pelepasannya dinamakan defekasi seluler. Bahan-bahan yang telah dicerna di dalam lisosom dapat kembali dilepaskan ke dalam sitoplasma dan selanjutnya terlibat di dalam proses katabolisme atau anabolisme.
C. Ultra Struktur dan Komposisi Lisosom
Lisosom merupakan vesikula berbatas membran tunggal, dimana di dalamnya terdapat enzim-enzim proteolitik. Membran lisosom mengandung karbohidrat netral, hexoamina, dan asan N-asetilmuramat yang lebih banyak dibandingkan dengan membran plasma.
Pengamatan dengan mikroskop elektron menunjukkan bahwa membran lisosom memiliki membran dengan tebal 9 nm, lebih tebal dari membran mitokondria. Membran lisosom memiliki kemampuan untuk berfusi secara selektif dengan membran sel yang lain, seperti fusi yang terjadi antara membran lisosom dengan fagosom atau endosom selama pencernaan intra sel. Demikian pula antara lisosom dengan membran plasma selama berlangsungnya sekresi sel.
Salah satu ciri lisosom adalah adanya kandungan berbagai enzim
hidrolase seperti fosfatase, nuklease, hidrolase, protease, dan
enzim-enzim untuk perombak lipida
| Jenis Enzim | Substrat | Asal Lisosom |
Fosfatase
ü RNA-ase ü DNA-ase Hidrolase v b- Galaktosidase v Glikosidase v Mannosidase v b- Glukuronidase v Lisosom v Hialuronidase v Asilsulfatase Protease Ø Katepsin Ø Kolagenase Ø Peptidase Enzim perombak lipida o Esterase o Fosfolipase |
ü RNA ü DNA v Galaktosidase v Glikogen v Mannosida v Polisakarida dan Mukopolisakarida v Dinding bakteri Mukopolisakarida v Asam hialuronat v kondritin sulfat v Sulfat organic Ø Protein Ø Kolagen Ø Peptida o Ester asam lemak o Fosfolipida |
ü Jaringan hewan, tumbuhan dan protista ü Jaringan hewan, tumbuhan dan protista v Jaringan hewan, tumbuhan v dan protista v Jaringan hewan v Jaringan hewan v Jaringan hewan v Ginjal v Hati v Hati, Tumbuhan Ø Sel hewan Ø Sel tulang Ø Jaringan hewan, tumbuhan dan protista o Jaringan hewan, tumbuhan dan protista o Tumbuhan |
Peranan Lisosom
Peranan fisiologi lisosom umumnya berhubungan dengan pencernaan intraseluler. Misalnya pencernaan makanan yang berlangsung pada protozoa dimana bahan-bahan yang berasal dari luar dicerna secara intraseluler atau heterofagi.
Endositosis merupakan persyaratan bagi pencernaan intraseluler bahan eksogen dengan molekul tinggi. Bukti menunjukkan bahwa vakuola makanan dihasilkan dari fusi antara endosom dengan lisosom primer.
Dalam darah, terdapat banyak sel-sel fagosit yang bekerja sebagai penghalang yang efektif dari invasi mikroorganisme atau benda-benda asing lainnya. Ada empat tipe fagosit darah, yaitu (i) Polymorpho Nuclear Neutrofic Leukocytes (PMNs), (ii) Eosinofil, (iii) Basofil, dan (iv) Monosit. Meskipun keempat tipe tersebut bersirkulasi di dalam darah, neutrofil dan manosit memiliki kemampuan untuk meninggalkan aliran darah dan mengembara di seluruh jaringan untuk menghilangkan bahan-bahan asing dalam jaringan dengan cara memfagositosisnya.
Monosit di dalam jaringan akan berkembang menjadi dewasa dan menjadi sel-sel makrofag. Berbagai jenis makrofag antara lain histiosit dalam jaringan pengikat, makrofag alveolar di dalam paru-paru, sel-sel kuffer di dalam jaringan hati, makrofag pelural di dalam peritoneal, osteoklas di dalam tulang, sel mikroglia di dalam sistim saraf pusat, sel schwann di dalam serabut saraf perifer, sel sinvial tipe A di dalam ruang sendi, dan makrofag di dalam jaringan limfoid dan jaringan ikat (Subowo, 1990).
Lisosom memainkan peranan yang sangat penting dalam resorbsi tulang yang dilakukan oleh osteoklas (gambar 7.10)
Selain itu, lisosom memegang peranan penting di dalam sekresi kelenjar tiroid oleh sel-sel epitel dari folikel tiroid.
kelenjar tiroid (Thorpe, 1984)
Lisosom memainkan peranan yang sangat penting selama berlangsungnya fertilisasi pada berbagai jenis hewan termasuk manusia, terutama selama berlang-sungnya reaksi akrosom. Enzim-enzim yang dilepaskan dari vesikula akrosom melakukan pencernaan terhadap selaput-selaput pelindung telur sehingga memungkinkan sel pronuklei jantan masuk menembus membran telur untuk berfusi dengan pronuklei betina .
badan golgi (Aparatus Golgi)

PENDAHULUAN
Badan golgi merupakan organel berbatas membran di dalam sel eukariota yang berperan dalam berbagai fungsi penting, meliputi (i) mengemas bahan-bahan sekresi yang dilepaskan dari sel, (ii) pemrosesan protein meliputi glikosilasi, fosforilasi, sulfasi dan proteolisis selektif, (iii) tempat utama sintesis karbohidrat, menyortir dan mendistribusikan produkproduk retikulum endoplasma (Sheeler dan Bianchii, 1983; Allar, 2005), (iv) sintesis glikolipida, dan (v)proliferasi elemen-elemen membran untuk membran plasma (Sheeler dan Bianchii, 1983)
Badan golgi ditemukan pada tahun 1898 oleh C. Golgi. Ia melaporkan adanya struktur-struktur internal yang dijumpai di dalam sel saraf yang olehnya diberi nama Apparato Reticular Interno atau struktur retikular internal (Thorpe, 1984). Atas dasar penemuan tersebut, pada tahun 1906, C, Golgi menerima hadiah nobel dalam bidang sitologi (Sheeler dan Bianchii, 1983). Kompleks golgi memiliki banyak nama, antara lain diktiosom, golgisom, lipokondria, idiosom, badan golgi, zat golgi, aparat golgi, kompleks golgi, dan vesikula golgi.
B. STRUKTUR BADAN GOLGI
Pengamatan dengan mikroskop elektron menunjukkan bahwa badan golgi tampak menyerupai kantung-kantung pipih dengan sejumlah struktur-struktur perifer yang bervariasi. Setiap kantung pipih diberi nama sakula atau lamella atau sisterna. Setiap sakula berbatas membran dengan tebal kurang lebih 7,5 nm dan di dalamnya terdapat ruang dengan lebar berkisar 15 nm yang diberi nama lumen (Sheeler dan Bianchii, 1983).
Pada sel tumbuhan, tumpukan sejumlah lamella dinama-kan diktiosom. Jarak antar lamella dalam suatu diktiosom berkisar 20 nm. Jumlah lamella pada suatu diktiosom kurang lebih 10 buah. Permukaan kompleks golgi yang terorientasi ke arah retikulum endoplasma disebut permukaan pembentukan atau permukaan cis. Sedang-kan permukaan yang lain disebut permukaan matang atau permukaan trans yang terorientasi ke arah membran plasma (Sheeler dan Bianchii, 1983).
Sisterna pada permukaan pembentukan berbentuk cembung, sedangkan sisterna pada permukaan matang berbentuk cekung. Vesikula-vesikula sederhana yang berada disekitar permukaan pembentukan akan berfusi dan berkontribusi menambah struktur badan golgi. Vesikulavesikula yang terdapat di sekitar permukaan matang lebih besar dan dibentuk dari permukaan sisterna. Vesikula-vesikula sederhana juga dilepaskan dari bagian tepi sisterna diantara permukaan pemebentukan dan permukaan matang.
Badan golgi pada kebanyakan sel terutama berfungsi dalam hubungannya dengan fungsi sekresi. Permukaan pembentukan yang terletak di dekat inti atau di dekat bagian khusus dari retikulum endoplasma yang tidak memiliki ribosom dinamakan retikulum endoplasma transisi. Membran inti dan retikulum endoplasma halus adalah sumber vesikula-vesikula sederhana yang berfusi dengan permukaan pembentukan. Beberapa vesikula-vesikula besar dibentuk dari permukaan matang dinamakan vesikula sekresi dan kelak akan berfusi dengan membran plasma. Jika vesikula-vesikula dilepaskan dari permukaan matang badan golgi, juga memungkinkan terbentuknya struktur-struktur internal sel seperti yang terjadi selama pembentukan akrosom pada sel sperma. Atau pembentukan lisosom (Sheler dan Bianchii, 1983.
B. LINTASAN SEKRESI
Secara morfologis, kompleks golgi menunjukkan suatu polaritas yang terkait secara langsung dengan fungsi organel. Di dalam sel-sel sekresi, kompleks golgi merupakan suatu pusat fungsional dalam rantai sekresi. Elemen-elemen penghubung lintasan sekresi tersebut adalah retikulum endoplasma, vesikula transisi, kompleks golgi, vesikula sekresi, dan membran plasma. Lintasan rantai sekresi ditunjukkan pada gambar 8.3.
Lintasan skretori meliputi minimal 6 tahap, yaitu (i) tahap sintesis, (ii) tahap segregasi, (iii) tahap transpor intraseluler, (iv) tahap konsentrasi, (v) tahap penyimpanan, dan (vi) tahap pengeluaran. Selama berlangsungnya sekresi, terjadi aliran materi dari retikulum endopla-sma ke arah badan golgi. Permukaan cekung badan golgi yang terorientasi ke arah retikulum endoplasma disebut permukaan pembentukan atau permukaan cis, sedangkan permukaan cembung yang terorientasi ke arah permukaan sel disebut permukaan matang atau permukaan trans.
Pada permukaan matang badan golgi, dibentuk vesikulavesikula sekresi, sedangkan pada permukaan pembentukan terdapat vesikula-vesikula transpor yang dibentuk dari membrane retikulum endoplasma. Selama berlangsungnya proses sekresi, terjadi aliran materi yang dibentuk dalam retikulum endoplasma dan bergerak melalui permukaan badan golgi dari permukaan pembentukan ke permukaan matang. Selanjutnya bergerak sebagai granula-granula sekresi pada daerah apikal sel dan pada akhirnya menuju ke membran plasma dimana bahanbahan tersebut dikeluarkan.
C. ASAL BADAN GOLGI
Menurut (Sheeler dan Bianchii, 1983) ada tiga sumber yang diusulkan yang diduga sebagai asal badan golgi, yaitu:
1. Vesikula-vesikula yang berasal dari membran luar salut inti atau retikulum endoplasma.
2. Vesikula-vesikula atau struktur-struktur sitoplasma yang lain.
3. Pembelahan dari badan golgi yang telah ada di dalam sel.
Sisterna dari golgi dapat dibentuk dari vesikula-vesikula yang berasal dari membran luar salut inti atau reticulum endoplasma. Vesikula-vesikula transisi bermigrasi ke permukaan pembentukan dari badan golgi dan selanjutnya berfusi dengan membran sisterna badan golgi yang sudah ada. Dengan cara ini, satu kompleks golgi dapat dibentuk secara sempurna.
Agregasi-agregasi dari vesikula transisi terjadi pada daerah sitoplasma yang disebut zona eksklusi (zones of exclusion) yang bebas ribosom. Zona tersebut biasanya dikelilingi oleh membran-membran retikulum endoplasma atau membran inti. Badan golgi-badan golgi sederhana diasumsikan telah ada pada awal perkembangan organel yang dijumpai pada zona tersebut. Beberapa bukti bahwa badan golgi terbentuk pada zona eksklusi (Sheeler dan Bianchii, 1983) adalah sebagai berikut:
1. Sel-sel pada biji yang dorman pada umumnya tidak memiliki badan golgi, namun pada zona eksklusi terdapat kumpulan vesikula-vesikula kecil. Hasil fotomikrograf pada sel biji pada stadium awal perkecambahan mendukung adanya per-kembangan badan golgi yang progresif pada zona eksklusi. Perkembangan badan golgi ber-tepatan dengan hilangnya vesikula-vesikula tran-sisi.
2. Pada telur katak, badan golgi tampak berkembang dari kelompok-kelompok vesikula yang terdapat pada zona eksklusi.
Selama berlangsungnya pembelahan sel pada sel hewan dan tumbuhan, jumlah badan golgi meningkat. Jumlah badan golgi yang dijumpai pada sel anak setelah pembelahan sama dengan jumlah badan golgi pada sel induk. Pada algae Botrydium granulatum yang multinukleat, pada setiap kutub sel yang sedang membelah tepat pada pembentukan spindel, terdapat sebuah badan golgi. Pada metafase lanjut, kedua badan golgi tersebut terdapat pada setiap ujung spindel dan dipisahkan oleh sentriol. Hal tersebut mendukung bahwa badan golgi dapat dibentuk melalui pembelahan organel.
Pembentukan badan golgi dari retikulum endoplasma melalui peleburan sisterna yang berbentuk tubular yang terdapat pada bagian perifer dari retikulum endop-lasma kasar. Hal ini terlihat pada sel-sel hati embrio. Pada saat sel-sel hati matang, sisterna tubular berubah menjadi bentuk mangkuk, kemudian bagian luar dari sisterna dibentuk vesikula-vesikula sekresi. Fenomena yang se-rupa dijumpai pada zoospora dan pembentukan sperma (gambar 7.4).
D. KOMPOSISI KIMIA BADAN GOLGI
Seperti halnya retikulum endoplasma, badan golgi juga mengandung senyawa lipida yang terdiri atas fosfolipida dan lemak netral. Sedangkan protein terdiri atas glikoprotein, mykoprotein, dan enzim.
1. Kandungan Lemak
Membran badan golgi memiliki kandungan lipida yang berbeda dari membran sel lain (tabel 8.1)
| Senyawa | % Fraksi Total | ||
| RE | BG | MP | |
| Total Fosfolipida Sfingomielin | 84,9 3,7 | 53,9 12,3 | 61,9 18,9 |
| Fosfatidilkolin Fosfatidilserin | 60,9 33,3 | 45,3 4,2 | 39,9 3,5 |
| Fosfatidilinositol | 8,9 | 8,7 | 7,5 |
| Fosfatidiletanolamin | 18,6 | 17,0 | 17,8 |
| Fosfatidilhialin | 4,7 | 5,9 | 6,7 |
| Lisofofatidiletanolamin | - | 6,3 | 5,7 |
| Total lipida netral | 15,1 | 46,1 | 38,1 |
| Kolesterol
Asam lemak bebas Trigliserida Ester-ester kolesterol |
|||
Analisa enzimatik dari badan golgi yang diisolasi didapatkan bahwa, badan golgi mengandung enzim-enzim yang sangat heterogen, misalnya enzim-enzim glikolisil transferase, oksidoreduktase, fosforase, dan sebagainya (tabel 8.2).
|
Seperti diungkapkan sebelumnya bahwa, fungsi dari kompleks golgi adalah sebagai tempat pemrosesan protein pasca translasi. Protein-protein tersebut berasal dari lumen retikulum endoplasma dan bergerak ke badan golgi. Ada dua cara bagaimana protein melewati permukaan pembentukan ke permukaan matang dari badan golgi, yaitu (i) Model Sisternal Progression atau model transport vesikula dan (ii) Model Sisternal Transfer atau model pematangan sisterna (Thorpe, 1984; Allar, 2005)
Pada model sisternal progression, vesikula-vesikula yang berisi protein yang berasal dari retikulum endoplasma berfusi dengan permukaan pembentukan dari badan golgi untuk mengalami proses lebih lanjut. Kemudian setiap sisterna bergerak melalui tumpukan badan golgi ke arah permukaan trans. Pada permukaan trans, sisterna dipecah-pecah menjadi sejumlah vesikula kecil yang membawa protein ke tujuannya. Kelemahan model ini adalah karena sisterna badan golgi mempunyai sifat-sifat yang berbeda, sehingga sulit untuk menjelaskan bagaimana satu sisterna dengan sifat-sifat tertentu dapat berubah menjadi sisterna dengan sifat-sifat lain (Thorpe, 1984; Allar, 2005).
Pada model sisternal transfer, protein bergerak dari satu sisterna ke sisterna yang lain melalui pembentukan vesikulavesikula kecil yang dilepaskan dari sisterna sebelumnya. Vesikula-vesikula tersebut bergerak maju dan berdifusi dengan sisterna berikutnya. (Thorpe, 1984; Allar, 2005)
Kompleks golgi berfungsi dalam biosintesis glikoprotein dan glikolipida. Glikoprotein adalah protein yang mengandung karbohidrat yang terikat secara kovalen, biasanya berupa Dgalaktosa, D-mannosa, L-fucosa, D-glukosamin, N-asetil-Dgalaktosamin, dan asam N-asetil-muramat atau asam sialat. Unit-unit monosakarida tersebut terikat dalam rantai oligosakarida.
Bahan-bahan yang akan disekresikan pada akhirnya berkumpul pada permukaan trans badan golgi dan kemudian dilepaskan dalam bentuk vesikula. Vesikula-vesikula sekresi melepaskan kandungannya dengan dua cara, yaitu secara konstitutif dan secara regulatif. Sejumlah protein-protein terlarut maupun yang terikat membran yang baru disintesis, lipida membran plasma yang baru disintesis dilepaskan dengan cara konstitutif, artinya tidak tergantung pada signal-signal tertentu seperti hormone atau neurotransmitter. Sejumlah proteinprotein tertentu yang tersimpan di dalam vesikula sekresi hanya dapat dilepaskan bilamana ia menerima sinyal-sinyal tertentu yang berasal dari hormone atau neurotransmitter. Sekresi seperti ini dinamakan sekresi regulative.
1. Sintesis, Pengemasan dan Pelepasan Hormon Peptida
Sintesis, pengemasan dan pelepasan hormon-hormon peptida melibatkan organel-organel sitoplasmik, yaitu reticulum endoplasma kasar bersama ribosom dan badan golgi. Tahap awal adalah sintesis protein pada yang berlangsung pada ribosom yang melekat pada retikulum endoplasma dan menghasilkan rantai polipeptida yang dikenal sebagai preprohormon. Rantai polipeptida tersebut diarahkan ke dalam lumen reticulum endoplasma oleh signal sequence asam-asam amino. Enzim-enzim pada retikulum endoplasma memotong signal sequence dan menghasilkan prohormon yang tidak aktif. Prohormon selanjutnya ditranspor menuju badan golgi. Di dalam badan golgi dikemas dan dilepaskan melalui pertunasan badan golgi dalam bentuk vesikula sekresi.
Di dalam vesikula sekresi terdapat sejumlah enzim yang berperan memotong prohormon menjadi satu atau lebih hormon yang aktif dan sejumlah fragemen-fragmen peptida. Hormon-hormon tersebut selanjutnya dirembeskan masuk ke dalam aliran darah untuk dibawah menuju jaringan target.
2. Sekresi Kelenjar Tiroid
Satu peranan kompleks golgi dalam biosintesis glikoprotein terlihat jelas pada sel-sel folikel tiroid. Sel-sel ini terlibat dalam sintesis tiroglobulin, suatu glikoprotein yang disekresikan oleh sel-sel ke dalam folikel dimana ia diiodinasi dan disimpan. Folikel adalah ruang interseluler yang besar dan dikelilingi oleh sel-sel epitel yang mensintesis tiroglobulin.
Tiroglobulin terdiri atas dua tipe rantai polipeptida yaitu tipe pertama yang hanya terdiri atas disakarida Nasetilglukosamin yang terikat pada mannosa dan tipe kedua yang lebih kompleks. Selain mengandung gula, juga mengandung galaktosa, fruktosa, dan asam sialat (Thorpe, 1984)
Urutan selama pembentukan tiroglobulin ditunjukkan pada gambar 8.6. Tiroglobulin diangkut secara vektorial dalam sel, dimulai pada retikulum endoplasma kasar dimana rantai polipeptida dibentuk. Polipetida mengalami prosesing berupa penambahan unit-unit monosakarida membentuk glikoprotein yang belum sempurna. Glikoprotein yang belum sempurna dibawa ke badan golgi melalui vesikula-vesikula transpor. Di dalam badan golgi glikoprotein mengalami penyempurnaan dengan penambahan unit-unit galaktosa (Thorpe, 1984).
Vesikula-vesikula sekresi yang mengandung glikoprotein yang telah sempurna dibentuk secara apikal dari kompleks golgi dan bergerak ke arah lumen folikel. Di dalam folikel berlangsung iodinasi. Glikoprotein yang telah diiodinasi kemudian disimpan dalam folikel-folikel hingga sel epitel distimulasi oleh TSH dari kelenjar pituitari untuk mengabsorbsi tiroglobulin. Hormon tiroid yang telah diiodinasi dipotong-potong dan selanjutnya dilepaskan ke dalam darah untuk selanjutnya menuju organ target.
Sel-sel folikel tiroid secara aktif mentranspor ion-ion iodine dari darah. Ion-ion tersebut selanjunya ditranspor menuju lumen folikel tiroid. Beberapa asam amino tirosin di dalam tiroglobulin akan mengalami iodinasi. Namun demikian ion-ion iodin tidak dapat terikat secara langsung pada asam amino tirosin. Oleh sebab itu ion-ion iodin harus melepaskan elektronnya melalui proses oksidasi. Oksidasi ion-ion iodin dikatalisis oleh enzimenzim peroksidase. 1 atau 2 ion-ion iodin yang telah mengalami oksidasi selanjutnya berikatan dengan asam amino tirosin membentuk T1 dan T2. T1 selanjuntnya bergandengan dengan T2 membentu T3 dan T2 bergandengan dengan T2 membentu T4.
Tiroglobulin yang telah mengalami iodinasi tetap berada di dalam folikel tiroid dalam bentuk koloid. Dibawah pengaruh hormone Thyroid stimulating hormon (TSH) yang berasal dari hipofisis, tiroglobulin selanjutnya dimasukkan di dalam sel-sel folikel tiroid secara endositosis. Di dalam sel-sel folikel, endosom yang mengandung tiroglobulin berfusi dengan lisosom primer dan selanjutnya berlangsung proses pemotongan. Pemotongan tiroglobulin tersebut oleh enzim-enzim lisosom menghasilkan hormone triiodotironin (T3) dan tetraiodotironin (T4). Selanjutnya hormone tersebut siap dirembeskan ke aliran darah.
3. Proliferasi membran-membran seluler
Kompleks golgi selain berperan dalam sekresi, juga memainkan peranan dalam persiapan protein-protein untuk organel-organel seperti lisosom dan membran plasma (gambar 8.8). Protein -protein yang dipersiapkan untuk lisosom atau membran plasma disintesis oleh ribosom-ribosom yang melekat pada retikulum endoplasma kasar. Beberapa dari proteinprotein tersebut dilepaskan ke dalam lumen reticulum endoplasma dan yang lain tetap pada membran reticulum endoplasma dan kelak menjadi dinding vesikula transpor. Dalam beberapa menit setelah sintesis, protein-protein tersebut tampak pada permukaan cis dari badan golgi.
Mekanise transport protein dari lumen RE ke badan golgi berlangsung melalui vesikula transport. Vesikula transport berfusi dengan permukaan cis badan golgi. Selanjutnya protein berpindah dari suatu sisterna ke sisterna berikutnya hingga mencapai permukaan trans. Protein-protein yang dipersiapkan untuk menjadi komponen membran lisosom dan membrane plasma tetap tertanam pada membrane RE. Protein tersebut diranspor dengan cara yang sama, namun pada saat tiba pada sasaran, protein tersebut tetap terikat pada membran.
4. Menyortir Protein-protein untuk Sasaran Tertentu
Protein-protein yang disintesis pada reticulum endoplasma yang diperuntukkan untuk membran plasma, lisosom, dan vesikula sekresi, diangkut ke badan golgi pada permukaan pembentukan. Protein-protein yang sampai pada permukaan pembentukan kompleks golgi bersama-sama dengan protein membran retikulum endoplasma. Vesikulavesikula yang dilepaskan dari permukaan trans tidak mengandung protein membran retikulum endoplasma. Kompleks golgi berperan memilih protein membran reticulum endoplasma oleh vesikula-vesikula kecil dari sisterna badan golgi permukaan pembentukan.
Rothman (1981 dalam Thorpe, 1984) membedakan permukaan pembentukan badan golgi dengan permukaan matang badan golgi. Permukaan pembentukan terdiri atas semua sisterna golgi kecuali satu atau dua yang terakhir. Peranan permukaan pembentukan adalah memilih protein retikulum endoplasma yang akan dikembalikan ke reticulum endoplasma. Permukaan matang dari badan golgi terdiri atas ½ sisterna permukaan yang berperan menerima protein yang telah dimurnikan dan menyebarkan melalui vesikula-vesikula ke lokasinya yang tepat di dalam sel. Rothman (1981) mengusulkan bahwa badan golgi terdiri atas tiga kompartemen yaitu kompartemen cis (kompartemen pembentukan), kompartemen medial, dan kompartemen trans (kompartemen matang). Kompartemen cis memilih dan melepaskan protein-protein retikulum endoplasma dan juga menambah gugus fosfat ke gula terminal protein lisosom. Kompartemen medial (terdiri atas sisterna di tengah-tengah tumpukan golgi) merupa-kan tempat penambahan Nasetilglukosamin. Sedangkan kompartemen trans merupakan tempat penambahan unit-unit galaktosa dan asam sialat, juga memilih berbagai protein sesuai dengan tujuan akhirnya. Penambahan gugus fosfat pada gula terminal protein lisosom dalam kompleks tersebut untuk mencegah penambahan Nasetilglukosamin dalam kompartemen medial dan penambahan galaktosa dan asam sialat dalam kompartemen trans.
5. Pembentukan Dinding Sel
Badan golgi berperan dalam pembentukan papan sel dan dinding sel. Papan sel dan dinding sel terbentuk selama anafase dan telofase mitosis dan miosis kedua. Sebelum anafase, kompleks golgi berada di luar kumparan. Selama anafase, kompleks golgi melepaskan vesikula-vesikula menuju pusat kumparan dan menimbun disekitar benang-benang kumparan. Vesikula berisi senyawa pembentuk papan sel dan dinding sel.
Vesikula-vesikula yang berasal dari badan golgi berisi bahan untuk pembentukan papan sel dan dinding sel. Di dalam sitoplasma terdapat mikrotubul-mikrotubul yang tersusun parallel dan disebut fragmoplas. Vesikula-vesikula yang ber asal dari badan golgi berhubungan dengan fragmoplas dan ditransportasikan sepanjang mikrotubul ke arah ekuatorial dan terakumulasi pada daerah dimana mikrotubul mengalami overlapping. Vesikula-vesikula yang mengandung bahan untuk papan sel dan dinding sel diakumulasikan pada daerah ekuatorial pada mikrotubul yang tumpang tindih dan berfusi membentuk papan sel. Bahan-bahan dari vesikula bergabung membentuk dinding sel. Vesikula-vesikula golgi yang baru terbentuk diakumulasi pada bagian tepi papan sel, kemudian berfusi dan meluas ke arah luar. Membran papan sel yang sedang merentang berfusi dengan membran plasma. Bahanbahan dinding sel dideposisikan membentuk dinding sel yang sempurna (Albert et al., 1983).
6. Pembentukan Akrosom
Akrosom adalah suatu badan berbatas membran yang terletak pada bagian kepala spermatozoa di sebelah anterior dari inti. Badan golgi merupakan organel yang berperan dalam pembentukan akrosom melalui beberapa fusi vesikula-vesikula yang mengandung bahan untuk akrosom. Sedangkan membrane vesikula-vesikula menjadi membran akrosom. Akrosom mengandung berbagai jenis enzim-enzim hidrolitik yang penting di dalam proses fertilisasi. Tanpa akrosom, maka sperma mengalami kesulitan untuk menembus berbagai selaput yang melindungi sel telur. Proses pembentukan akrosom pada kepala sperma.
E. tetanus
No comments:
Post a Comment