OPO TO LEMAK
Yamg jelas orang berlemak pasti disebut Gendut ...........O jadi lemak bikin gendut
- Lemak yang tidak diperlukan disimpan di dalam Jaringan adipose (bawah kulit). Ia dikeluarkan apabila diperlukan dan diubah menjadi gliserol dan asam lemak di dalam hati.
- Apabila lemak telah di metabolismekan oleh hati, terdapat zat sisa berupa keton
- Lemak atau lipid terdapat pada semua bagian tubuh manusia terutama pada bagian otak, mempunyai peran yang sangat penting dalam proses metabolisme secara umum. Sebagian lipid jaringan tersebar sebagai komponen utama membrane sel dan berperan mengatur jalannya metabolisme di dalam sel.
- Komponen struktur membran

- Lapisan pelindung pada beberapa jasad
- Bentuk energi cadangan
- Komponen permukaan sel yang berperan dalam proses interaksi antara sel dengan senyawa kimia di luar sel, seperti dalam proses kekebalan jaringan
- Sebagai komponen dalam proses pengangkutan melalui membran.
- Biosintesis asam lemak sebagai bagian dari biosintesis lipida adalah suatu proses metabolisme yang penting
- Hal ini benar jika diingat jaringan hewan mempunyai kemampuan terbatas untuk menyimpan energi dalam bentuk karbohidrat, dalam hal ini sebagian dari polisakarida dirombak melalui proses glikolisis menjadi asetil ko-A, yang merupakan pra zat ( zat antara) untuk biosintesis asam lemak dan triasilgliserol.
- Senyawa lipid ini mempunyai kandungan energi yang lebih tinggi (1 gr = 9,3 kal) bila dibandingkan dengan karbohidrat ( 1 gr = 4,1 kal) dan dapat disimpan sebagai cadangan energi yang besar di dalam jaringan lemak ( Adiposa)
- Di dalam tumbuhan, senyawa lipid disimpan sebagai cadangan energi yang cukup besar di dalam biji dan buah.
- Biosintesis asam lemak dari asetil ko-A terjadi di hampir semua bagian tubuh hewan, terutama di dalam jaringan hati, jaringan lemak dan kelenjar susu.
- Biosintesis ini berlangsung dalam sitoplasma, membutuhkan asam sitrat sebagai kofaktor dan membutuhkan CO2 sebagai factor pembantu dalam mekanisme pemanjangan rantai asam lemak, meskipun CO2 tidak tergabung ke dalam asam lemak tersebut.
- Asam lemak adalah suatu senyawa yang terdiri dari rantai panjang hidrokarbon dan gugus karboksilat yang terikat pada ujungnya.
- Pertama, sebagai satuan pembentuk fosfolipid dan glikolipid yang merupakan molekul amfipatik sebagai komponen membran biologi.
- Asam lemak mempunyai peran yang sangat penting sebagai sumber pembentuk energi dalam tumbuhan dan hewan.
- Sebagian besar asam lemak disimpan dalam bentuk senyawa trigliserida di dalam sel.
- Sebagian besar asam lemak bebas yang mengalami katabolisme berasal dari proses hidrolisis trigliserida oleh enzim lipase yang terdapat di dalam sel jaringan lemak.
- Asam lemak ini dikeluarkan dari sel, berikatan dengan serum albumin yang kemudian bersama aliran darah dibawa ke jaringan lainnya di dalam tubuh untuk selanjutnya mengalami oksidasi.
- Dalam hal ini asam lemak yang masuk ke jaringan lebih dulu dipergiat dengan perantaraan enzim di dalam sitoplasma, baru kemudian dapat dimasukkan ke dalam mitokondrion untuk selanjutnya mengalami proses oksidasi menghasilkan energi yang dipakai untuk segala kegiatan dalam tubuh yang memerlukan energi. OK
- Oksidasi sempurna asam lemak berantai panjang di dalam semua sel jaringan hewan mamalia, kecuali di dalam sel otak, menghasilkan CO2 dan H2O sebagai hasil akhir.
- Dalam keadaan tertentu oksidasi asam lemak dalam sel otak menghasilkan asam β-hidroksibutirat.
- Kelincahan gerak, penyebaran, dan oksidasi asam lemak yang terjadi di dalam tubuh berlangsung secara terpadu dengan proses metabolisme karbohidrat dan diatur oleh sistem hormon endokrin yang rumit.
 OKSIDASI ASAM LEMAK
OKSIDASI ASAM LEMAK
- Asam lemak yang ada di dalam tubuh banyak mengalami oksidasi dalam β-oksidasi menjadi asetil KoA.
- Oksidasi asam lemak ini terjadi di dalam mitokondria.
- Untuk memasuki mitokondria, asam-asam lemak pertama-tama harus diubah menjadi suatu bentuk asil-KoA oleh aksi tiokinase dan ATP dalam mikrosom atau pada permukaan mitokondria.
- Untuk asam-asam lemak rantai panjang, biasanya harus diubah terlebih dahulu menjadi asilkarnitin supaya dapat masuk menembus membran mitokondria.
- Sesampainya di dalam mitokondria, barulah asam lemak dapat dioksidasi.
- Semua proses ini mulai dari masuk ke dalam mitokondria hingga mengalami oksidasi terjadi dalam 3 tahap.
- Aktivasi asam lemak yang terjadi di sitoplasma
- Transport asam lemak ke dalam mitokondria
- Proses β-oksidasi di dalam matriks mitokondria
- Asam lemak diaktifkan menjadi bentuk asil KoA oleh tiokinase atau Asil KoA sintetase (tiokinase).
- Reaksi ini terjadi dalam dua langkah dan membutuhkan ATP, KoA dan Mg2+. Asam lemak bereaksi dengan ATP membentuk asiladenilat yang kemudian bergabung dengan KoA untuk menghasilkan asil KoA.
- Dalam proses aktivasi ini dibutuhkan 2 fosfat berenergi tinggi karena ATP akan diubah menjadi pirofosfat (PPi).
- Enzim inorganik pirofosfatase menghidrolisis PPi menjadi fosfat (Pi) dan proses ini bersifat irreversibel.
- Aktivasi asam lemak ini umumnya terjadi di retikulum endoplasma, peroksisom dan membran luar mitokondria.
- Sebab pada daerah-daerah ditemukan enzim asil KoA sintetase yang berfungsi mengaktifkan asam lemak.
- Dalam aktivasi asam lemak ada yang harus diperhatikan. Yaitu panjang rantai asam lemak yang akan diaktivaskan.
- Apakah asam lemak tersebut berantai panjang (10-20 karbon), medium (4-12 karbon) ataupun berantai pendek (<4>
Transport asil KoA ke dalam mitokondria
- Asil KoA (asam lemak yang sudah diaktifkan) yang berantai panjang tidak dapat menembus membran mitokondria dengan mudah.
- Oleh karena itu, Asil KoA berantai panjang akan diubah dulu menjadi asilkarnitin oleh karnitin parmitoiltransferase-I (karnitin asiltransferase-I) yang ada di sitoplasma agar dapat masuk ke dalam mitokondria.
- Semua proses ini terjadi dalam 4 tahap
- Grup Asil pada asilKoA ditransfer ke karnitin yang dibantu oleh karnitin asiltransferase-I. Pada tahap ini asilKoA akan diubah menjadi Asilkarnitin.
- Asilkarnitin akan dibawa masuk menembus membran mitokondria sampai ke matriks oleh protein carrier.
- Karnitin asiltransferase-II (yang berada di membran dalam mitokondria) akan mengkonversi asilkarnitin menjadi asilKoA kembali sedangkan karnitin akan dilepas.
- Karnitin akan dilepaskan ke sitoplasma untuk digunakan kembali.
- Setiap siklus β-oksidasi akan membebaskan dua unit karbon asetil KoA dan terjadi dalam 4 urutan reaksi.
- Oksidasi: Asil KoA mengalami dehidrogenasi oleh FAD-dependent flavoenzim, asilKoA dehidrogenase. Ikatan double dibentuk antara karbon α dan β.
- Hidrasi: Enoil KoA hidratase menghidrasi ikatan double tadi sehingga membentuk β-hidroksiasil KoA.
- Oksidasi: Dalam tahap ini akan dihasilkan β-ketoasil KoA.
- Pemecahan (Cleavage): Reaksi terakhir β-oksidasi akan membebaskan 2 karbon asetil KoA dari asil KoA.
- Pada jalur β-oksidasi, asam lemak yang jumlah atom karbonnya ganjil, akan membentuk asetil KoA hingga tersisa sebuah residu tiga karbon (propionil KoA). Propionil KoA ini akan diubah menjadi suksinil KoA yang siap memasuki siklus asam sitrat.
 Pada oksidasi asam lemak, asam lemak akan diubah dalam proses β-oksidasi menjadi asetil KoA.
Pada oksidasi asam lemak, asam lemak akan diubah dalam proses β-oksidasi menjadi asetil KoA.- Proses Oksidasi lain dengan α-oksidasi
- Meskipun β-oksidasi merupakan jalur paling dominan pada oksidasi lemak, namun sebenarnya masih ada jalur oksidasi lain yang dikenal, yaitu α-oksidasi. α-oksidasi melakukan penghapusan/penghilangan satu atom karbon pada asam lemak dan tidak melibatkan ikatan asam lemak dengan koenzim A.
- Dalam proses ini tidak ada energi apapun yang dihasilkan.
- Sebelum memulai oksidasi, terlebih dahulu asam lemak dihidroksilasi. Baik itu hidroksilasi pada asam lemak rantai pendek maupun hidroksilasi asam lemak rantai panjang (untuk sintesis sfingolipid).
- Proses hidroksilasi ini kemungkinan besar terjadi di retikulum endoplasma dan mitokondria yang melibatkan “fungsi oksidasi campuran.”
- Proses Oksidasi lain dengan ω-oksidasi
- Jalur ini termasuk jalur yang jarang. jalur ini terjadi di retikulum endoplasma pada hampir semua jaringan tubuh.
- Sama seperti α-oksidasi, ω-oksidasi juga melibatkan hidroksilasi sebelum oksidasi asam lemak.
- Dalam hal ini hidroksilasi terjadi pada karbon metil di akhir gugus karboksil atau karbon disebelah metil akhir.
- Jalur ini juga melibatkan “fungsi oksidasi campuran” (mixed function oxidase) dan membutuhkan sitokrom P450, O2, dan NADPH.

- Bahan dari piruvat membuat proses ini seolah-olah lebih mudah.
- Piruvat diubah menjadi asetilKoA dan oksaloasetat, yang akhirnya akan bergabung kembali membentuk sitrat.
- Karbohidrat dan asam amino yang dikonsumsi berlebihan akan dikonversi menjadi asam lemak dan disimpan sebagai triasilgliserol.
- Dan proses ini (selanjutnya kita sebut sintesis asam lemak) paling banyak terjadi di hati, ginjal, jaringan adiposa dan kelenjar mamaria.
- Dalam proses ini, asetil KoA bertindak sebagai substrat langsung atau bahan utamanya, sedangkan palmitat sebagai produk akhirnya.
- Sintesis asam lemak melibatkan asetil KoA dan NADPH. Asetil KoA disini berfungsi sebagai sumber atom karbon sementara NADPH berperan sebagai bahan pendukungnya saja.
Sintesis asam lemak terjadi dalam 3 proses. Yang diantaranya:
- Produksi asetil KoA dan NADPH
- Pembentukan Malonil KoA dari asetil KoA
- Reaksi kompleks sintesis asam lemak

Produksi asetil KoA dan NADPH
- Asetil KoA dan NADPH merupakan syarat paling penting dalam sintesis asam lemak.
- Asetil KoA diproduksi di dalam mitokondria melalui oksidasi asam lemak dan piruvat, asam amino dan juga dari badan keton.
- Seperti yang sudah di atas sebelumnya, bagaimana oksidasi asam lemak dapat menyediakan asetil KoA di dalam mitokondria.

- Dimulai dari proses yang terjadi di sitoplasma sampai ke dalam mitokondria.
- Asetil KoA yang dihasilkan tersebutlah yang menjadi salah satu sumber bahan untuk sintesis asam lemak ini.
- Sedangkan sumber asetil KoA yang diperoleh dari piruvat disediakan oleh piruvat dehidrogenase.
- Piruvat yang masuk ke dalam mitokondria akan diubah menjadi asetil KoA dan oksaloasetat.
- Piruvat dehidrogenase akan merubah piruvat menjadi asetil KoA sedangkan piruvat karboksilase mengubah piruvat menjadi oksaloasetat.
- Sedangkan bahan NADPH dapat diperoleh dari jalur pentosa fosfat dan bisa juga dari NADPH yang dihasilkan enzim malat.
- Kemudian, untuk memulai proses sintesis asam lemak, asetil KoA akan bergabung terlebih dahulu dengan oksaloasetat membentuk sitrat.
- Asetil KoA harus diubah dulu menjadi sitrat karena asetil KoA tidak mampu menembus membran mitokondria.

- Sitrat yang baru saja dibentuk mampu dengan bebas menembus membran mitokondria sampai ke sitoplasma.
- Di sitoplasma sitrat ini akan dipecah oleh sitrat liase menjadi asetil KoA dan oksaloasetat.
- Pada tahap ini, oksaloasetat diteruskan hingga membentuk malat sedangkan asetil KoA dilanjutkan ke proses berikutnya, yaitu pembentukan malonil KoA dari asetil KoA.

Comparison of Energy Yields and Oxygen Consumption for Fats vs. Ketones vs. Carbohydrates:
1 NADH = 3 ATP
1 FADH2 = 2 ATP
Palmitate (3 molecules = 48 "C"s)
- Step 1: b-oxidation to acetyl CoA (6 cycles); (1 NADH + 1 FADH2)/cycle Step 1cont'd: 7 x 3 molecules 21 FADH2+ 21 NADH ==> +105 ATP; -42 O atoms
- Step 2: Acetyl CoA oxidation via TCA cycle; (3 NADH, 1 FADH2, 1 GTP) / Acetyl CoA Step 2 cont'd: 8(6 + 2) x 3 molecules: 24 Acetyl CoA ==> +288 ATP; -96 O atoms
- Step 3: Acyl CoA formation (ATP --> AMP + PPi; 2 ATP / molecule) Step 3 cont'd: 2 x 3 molecules: ==> -6 ATP
b-hydroxybutyrate (12 molecules = 48 "C"s)
- Step 1: oxidation to acetoacetate via b-hydroxybutyrate DH Step 1 cont'd: 1 x 12 molecules: 12 NADH ==> +36 ATP; -12 O atoms
- Step 2: Acetoacetate cleaved to 2 acetyl CoA ( loss 1 GTP due to succinyl CoA diversion) Step 2 cont'd: 1 x 12 molecules: -12 GTP ==> -12 ATP
- Step 3: Oxidation of acetyl CoA via TCA cycle Step 3 cont'd: 2 x 12 molecules: 24 Acetyl CoA ==> +288 ATP; -96 O atoms
Glucose (8 molecules = 48 "C"s)
- Step 1: Aerobic glycolysis, 2 NADH (mal-asp shuttle) + 2 ATP/glucose Step 1 cont'd: 2 x 8 molecules: 16 NADH + 16 ATP ==> +64 ATP; -16 O atoms
- Step2: PDH Step 2 cont'd: 2 x 8 molecules: 16 NADH ==> +48 ATP; -16 O atoms
- Step 3: Oxidation of acetyl CoA via TCA cycle Step 3 cont'd: 2 x 8 molecules: 16 Acetyl CoA ==> +192 ATP; -64 O atoms
- Asetil KoA dikarboksilasi menjadi malonil KoA oleh asetil KoA karboksilase.
- Malonil KoA nantinya akan mendonor 2 unit karbon untuk ditambahkan ke rantai asam lemak yang sedang tumbuh pada kompleks asam lemak sintase.
- Proses pembentukan ini membutuhkan vitamin biotin.
- karboksilasi biotin yang membutuhkan ATP dan
- pembentukan malonil KoA dengan pemindahan gugus karboksil ke asetil KoA.

- Saat asetilKoA karboksilase diaktifkan kadar malonil KoA akan meningkat.
- Saat sintesis asam lemak berlangsung, malonil KoA akan menginhibisi oksidasi asam lemak agar asam lemak yang akan terbentuk nantinya tidak langsung dioksidasi.
- Asam lemak sintase merupakan enzim besar yang terdiri dari dimer yang identik, yang masing-masing subunitnya (monomer) memiliki tujuh aktivitas enzim asam lemak sintase pada rantai polipeptida.
- Setiap monomernya berberat molekul 240.000 dan memiliki sebuah protein pembawa asil (ACP, acyl carrier protein).
- Fungsi ACP dalam sintesis asam lemak adalah bertindak sebagai suatu karier perantara. Segmen ACP memiliki sebuah residu 4- fosfopanteteinil yang berasal dari pemutusan koenzim A.
- Kedua subunit tersebut tersusun (kepala ke leher).
- Salah satu subunit bergandengan dengan gugus fosfopanteteinil sulfhidril sedangkan subunit yang lainnya bergandengan dengan sisteinil sulfhidril.
- Pada proses ini, gugus asetil dari asetil KoA akan dipindahkan ke gugus fosfopanteteinil sulfhidril ACP pada satu subunit, dan kemudian ke gugus siteinil sulfhidril pada subunit yang lainnya.
- Gugus malonil dari malonil Ko A kemudian melekat ke gugus fosfopanteteinil sulfhidril ACP pada subunit pertama.
- Gugus asetil dan malonil berkondensasi sehingga menyebabkan pelepasangugus karboksil malonil sebagai karbondioksida. Kemudian sebuah rantai α-keto asil (C4) akan melekat pada gugus fosfopanteteinil sulfhidril.
- Malonil KoA mampu menghambat kerja oksidasi lemak dengan menginhibisi Karntinasiltransferase-I yang berperan membawa asam lemak rantai panjang ke dalam mitokondria dari sitoplasma. Secara otomatis, asam lemak gagal dibawa masuk ke dalam mitokondria sehingga oksidasi asam lemak tidak terjadi.Mekanisme ini berfungsi untuk mencegah asam lemak yang baru terbentuk langsung mengalami oksidasi.
- Pemindahan gugus asetil dari asetil KoA ke ACP dikatalisis oleh enzim asetil KoA-ACP transasilase. Sedangkan pemindahan gugus malonil dari malonil KoA ke ACP dibantu oleh enzim malonil KoA-ACP transasilase
- Gugus malonil (dari malonil KoA) dan gugus asetil (dari asetil KoA) melekat pada gugus fosfopanteteinil sulfhidril ACPRantai asil lemak 4-karbon tersebut kemudian dipindahkan\ ke gugus sisteinil sulfhidril dan kemudian bergaung dengan sebuah gugus malonil.
- Urutan reaksi ini terus menerus dilakukan sehingga panjang rantai mencapai 16 karbon (palmitat).
- Dalam tahap ini, palmitat dibebaskan. Selanjutnya palmitat dapat mengalami desaturasi atau pemanjangan rantai.
- Asetoasetil KoA merupakan kondensasi antara dua molekul asetil KoA
- sedangkan HMG KoA merupakan gabungan antara asetoasetil KoA dan satu molekul asetil KoA.
- Asetoasetat, β-hidroksibutirat (D-3-hidroksibutirat), dan aseton merupakan senyawa-senyawa keton yang sangat penting bagi tubuh.
- Apabila laju oksidasi asam lemak tinggi, hati akan memproduksi banyak asetoasetat dan β-hidroksibutirat.
- Proses ketogenesis ini terjadi di dalam matriks mitokondria dengan asetil KoA sebagai bahan utamanya.
- Asetil KoA yang dibentuk dari oksidasi asam lemak, piruvat, atau beberapa asam amino merupakanBprekursor badan keton.
- Proses ketogenesis terjadi melalui tahap-tahap berikut:
- Dua mol asetil KoA hasil β-oksidasi bergabung dan membentuk asetoasetil KoA yang dikatalisis oleh enzim tiolase.
- Asetoasetil KoA yang baru saja terbentuk akan bergabung dengan molekul asetil KoA yang lain untuk membentuk β-hidroksi β-metil glutaril BKoA (HMG-KoA).
- HMG-KoA dipecah menjadi asetoasetat dan melepas asetil KoA oleh HMG-KoA liase.
- Asetoasetat secara spontan dapat mengalami dekarboksilasi sehingga membentuk aseton yang termasuk salah satu senyawa keton.
- Kemudian asetoasetat juga dapat tereduksi menjadi β-hidroksibutirat.
 Asetoasetil KoA merupakan kondensasi antara dua molekul asetil KoA sedangkan HMG KoA merupakan gabungan antara asetoasetil KoA dan satu molekul asetil KoA
Asetoasetil KoA merupakan kondensasi antara dua molekul asetil KoA sedangkan HMG KoA merupakan gabungan antara asetoasetil KoA dan satu molekul asetil KoA SINTESIS TRIASILGLISEROL
- Sintesis triasilgliserol paling sering terjadi di hati dan di sel lemak.
- Triasilgliserol merupakan ester dari gliserol dan asam lemak.
- Di hati gliserol 3 fosfat dapat diperoleh dari fosforilasi gliserol dan dari glikolisis.
- Gliserol yang ada di hati difosforilasi oleh enzim gliserol kinase.
- Sayangnya jaringan adiposa tidak memiliki enzim gliserol kinase ini sehingga pasokan gliserol 3 fosfat di jaringan adiposa hanya diperoleh dari jalur glikolisis. OK


Pada gambar diatas, jalur glikolisis dimulai dari bahan glukosa hingga menjadi bentuk DHAP (Dalam gambar tersebut jalur glikolisis hanya ditampilkan secara singkat, tidak dipaparkan secara jelas).
Dihidroksiaseton fosfat (DHAP) selanjutnya direduksi oleh gliserol 3 fosfat
Dehidrogenase menjadi gliserol 3 fosfat.
- Proses selanjutnya dapat diterangkan dengan tahap-tahap berikut:
- Gliserol 3-fosfat yang sudah tersedia (baik dari fosforilasi gliserol maupun dari jalur glikolisis) akan ditambahkan dengan grup asil.
- Proses ini dikatalisis oleh gliserol 3-fosfat asiltransferase sehingga akan membentuk asam lysofosfatidat.
- Grup asil lainnya akan ditambahkan pada asam lysofosfatdat untuk membentuk asam fosfatidat. Proses ini juga dikatalisis oleh enzim asiltransferase. Asam fosfatidat mengalami defosforilasi danmenghasilkan diasilgliserol.
- Diasilgliserol bergabung dengan grup asil yang lain yang dikatalisis oleh asiltransferase hingga membentuk triasilgliserol.
- Tiga asam lemak yang ditemukan di triasilgliserol bukanlah asam lemak yang sama.
- Pada karbon 1 ditemukan asam lemak jenuh (misal asam palmitat)
- sedangkan pada karbon 2 dan 3 dapat ditemukan asam lemak tidak jenuh (misal asam oleat).
Proses biosintesis kolesterol dapat dijelaskan dalam beberapa tahap berikut.

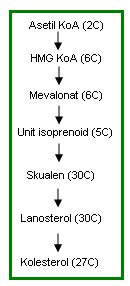
- Sintesis HMG KoA (β-hidroksi β-metilglutaril KoA) Proses ini mirip dengan proses pembentukan HMG KoA dalam mekanisme ketogenesis. Hanya berbeda lokasi saja. Ketogenesis terjadi di mitokondria
- sedangkan sintesis kolesterol terjadi di sitoplasma.
- terjadi di dalam mitokondria (ketogenesis) dan
- terjadi di sitoplasma (sintesis kolesterol).
- Dua molekul asetil KoA awalnya berkondensasi membentuk asetoasetil KoA.
- Dan molekul asetil KoA lainnya ditambahkan sehingga menghasilkan HMG KoA.
- Enzim HMG KoA akan mereduksi HMG KoA menjadi mevalonat.
- Enzim ini berada di retikulum endoplasma.
- Pada proses reduksi ini dibutuhkan ekivalen pereduksi yang disuplai oleh NADPH. HMG KoA juga sangat dipengaruhi oleh hormon misalnya seperti hormon insulin dan glukagon.
- Jika kadar glukagon meningkat, HMG KoA reduktase mengalami fosforilasi dan menjadi tidak aktif sedangkan jika kadar insulin yang meningkat, enzim tersebut akan mengalami defosforilasi dan menjadi aktif.
- Begitu juga dengan hormon tiroid dan glukokortikoid. Hormon tiroid meningkatkan aktivitas reduktase ini sedangkan glukokortikoid menurunkannya.
- Selanjutnya mevalonat mengalami fosforilasi oleh ATP. Kemudian mevalonat kinase mengkonversi mevalonat menjadi 3-fosfo 5-pirofosfomevalonat. Yang terjadi berikutnya adalah dekarboksilasi oleh enzim fosfomevalonat sehingga terbentuk isopentenil pirofosfat.
- Kesimpulannya, reaksi pertama dari tahap ini adalah mevalonat diubah menjadi 3-fosfo 5- pirofosfomevalonat yang kemudian didekarboksilasi menjadi isopentenil pirofosfat.
- Isopentenil pirofosfat inilah salah satu unit isoprenoid yang dimaksud. Sedangkan unit isoprenoid lainnya adalah 3,3-dimetilali pirofosfat. 3,3-dimetilali pirofosfat diperoleh dari reaksi isomerase yang dikatalisis oleh enzim isomerase isopentenil pirofosfat. Dalam beberapa referensi, unit isoprenoid disebut unit 5-karbon isoprenoid.
- Isopentenil pirofosfat (Isopentenyl pyrophosphate, IPP) dan 3,3-dimetilalil pirofosfat (dimethylallyl pyrophosphate, DPP) berkondensasi membentuk geranil pirofosfat (10C).
- Dalam tahap ini terjadi penambahan satu unit isoprenoid lagi untuk menghasilkan farnesil pirofosfat.
- Unit isoprenoid ditambahkan disini adalah satu molekul IPP. Molekul IPP tersebut akan berkondensasi dengan GPP untuk membentuk farnesil pirofosfat (15C).
- Dua unit farnesil pirofosfat bergabung dan direduksi sehingga menghasilkan skualen (30C).
- Pada reaksi selanjutnya enzim skualen monooksigenase mengubah skualen menjadi skualen 2,3 epoksida.
- Reaksi ini membutuhkan NADPH dan oksigen molekular (O2). Kemudian skualen 2,3 epoksida mengalami siklisasi untuk menghasilkan lanosterol.
- Reduksi atom karbon dari 30C menjadi 27C.
- Penghilangan dua gugus metil dari C4 dan satu gugus metil dari C14.
- Pemindahan ikatan rangkap dari C8 ke C5.
- Reduksi ikatan rangkap antara C24 dan C25.
- etelah keempat reaksi penting di atas selesai, kolesterol akhirnya terbentuk.

Kandungan lemak yang terbanyak pada adiposit adalah trigliserida dan kolesteril ester. Sekresi yang dihasilkan antara lain resistin, adiponektin, leptin, estradiol, angiotensin dan sitokina, interleukin.
Meskipun adiposit dapat dihasilkan dari fibroblast maupun sel punca mesenchymal, asal-muasal adiposit yang menyusun jaringan adiposa masih belum dapat diketahui dengan pasti.
Adiposit menghasilkan dan mensekresi beberapa protein yang berperan sebagai hormon. Hormon yang dikenal
sebagai adiponektin, berperan penting dalam proses radang, dan aterosklerotik. Adiponektin merupakan salah satu dari banyak faktor spesifik jaringan adiposa. Adiponektin berperan memperbaiki sensitivitas insulin dan menghambat peradangan vaskuler. Kadar adiponektin di dalam plasma secara bermakna menurun pada subyek yang mengalami obesitas, resistensi
Lemak adalah topik diskusi sehari-hari di negeri ini. Kita memahami ini karena dapat membawa risiko kesehatan yang luar biasa dan bisa menghancurkan tubuh kita, tapi yang mengejutkan, adalah hal ini penting untuk fungsi sehari-hari. Dalam pelajaran ini, kita akan menjelajahi jawaban di balik jaringan adiposa tubuh - jaringan ikat yang mengandung lemak - dan melihat bagaimana melindungi, menghangatkan, dan bahan bakar kehidupan kita sehari-hari.
Jaringan adiposa (lemak tubuh) sangat penting untuk
kesehatan. Seiring dengan sel-sel lemak, jaringan adiposa mengandung sel-sel
saraf banyak dan pembuluh darah, menyimpan dan melepaskan energi untuk bahan
bakar tubuh dan melepaskan hormon-hormon penting penting untuk kebutuhan tubuh.
Nama alternatif untuk jaringan adiposa
Lemak, lemak tubuh.
Dimana jaringan adiposa saya?
Jaringan adiposa umumnya dikenal sebagai lemak tubuh. Hal
ini ditemukan di seluruh tubuh. Hal ini dapat ditemukan di bawah kulit (lemak
subkutan), dikemas di sekitar organ internal (lemak visceral), antara otot,
dalam sumsum tulang dan jaringan payudara. Pria cenderung menyimpan lemak
visceral lebih (sekitar organ internal mereka), yang mengarah ke obesitas
sekitar pertengahan perut mereka. Namun, perempuan cenderung menyimpan lebih
banyak lemak subkutan dalam bokong dan paha. Perbedaan ini karena hormon seks
yang diproduksi oleh pria dan wanita.
Apa yang jaringan adiposa lakukan?
Jaringan adiposa sekarang dikenal sebagai organ endokrin
yang sangat penting dan aktif. Hal ini juga ditetapkan bahwa adiposit, (atau
sel-sel lemak), memainkan peran penting dalam penyimpanan dan pelepasan energi
di seluruh tubuh manusia. Baru-baru ini, fungsi endokrin adiposa telah
ditemukan. Selain adiposit, jaringan adiposa mengandung banyak sel-sel lain
yang mampu memproduksi hormon tertentu dalam merespon sinyal dari sisa organ di
seluruh tubuh. Melalui tindakan ini hormon ', jaringan adiposa memainkan peran
penting dalam regulasi glukosa, kolesterol dan metabolisme hormon seks.
Peran Fungsi Pentingnya adiposa
Bayangkan ada dua orang yang saling pukul diatas kolam
dalam permainan memeriahkan hari kemerdekaan, yang satu kurus badannya
sedangkan yang lainnya berbadan gemuk.
Mana yang akan lebih baik selama pertempuran? Siapakan
yang akan mengalami memar lebih lama? Ini mungkin tampak sulit untuk percaya,
tetapi seseorang dengan lebih banyak lemak dalam tubuhnya tidak akan terlalu
sakit dan memar pada hari berikutnya. Fungsi jaringan adiposa sebagai bantalan
terhadap cedera fisik bagi jaringan tubuh. Banyak organ utama yang dibungkus
dalam lapisan lemak viseral, jauh di dalam, untuk melindungi organ selama
trauma fisik.
Manakah dari mereka yang bertarung akan memiliki kontrol
yang lebih baik atas suhu tubuhnya selama permainan? Sekali lagi, orang yang
memiliki persentase lemak tinggi akan mampu mempertahankan suhu inti tubuh
tanpa harus memakai lapisan besar pakaian hangat. Fungsi jaringan adiposa
sebagai isolator panas alami.
Lemak merupakan salah satu sumber utama energi tubuh.
Makanan yang dimakan dan tidak dibakar untuk bahan bakar langsung disimpan
sebagai lemak dalam sel lemak. Hal ini dapat dikonversi menjadi bahan bakar
ketika tubuh kehabisan sumber energi langsung dari bahan karbohidrat. Mengubah
lemak menjadi bahan bakar yang dapat digunakan datang dengan biaya tinggi, dan
tubuh harus mengeluarkan dua kali lipat energi untuk mengubahnya menjadi bahan
bakar dibandingkan dengan karbohidrat atau protein. Oleh karena itu, otak
umumnya akan mengerahkan segala daya pilihan lainnya karbohidrat dan sumber
protein pertama. Lemak yang dibakar untuk bahan bakar memiliki waktu konversi panjang,
yang berarti aktivitas harus dipertahankan untuk jangka waktu yang panjang. Dan
bahan bakar membutuhkan pasokan oksigen yang sangat tinggi untuk mengubahnya.
Oleh karena itu sedikit lemak yang terbakar selama latihan keras selama periode
waktu yang singkat (seperti pertandingan sepak bola). Jika anda ingin membakar
lemak, anda akan lebih baik pergi untuk berjalan-jalan yang lama daripada,
pertandingan sepak bola halaman belakang dengan waktu yang pendek.
Hormon apa yang diproduksi jaringan adiposa?
Sejumlah hormon yang berbeda dilepaskan dari jaringan
adiposa dan ini bertanggung jawab untuk fungsi yang berbeda dalam tubuh. Contoh
ini adalah:
Aromatase yang
terlibat dalam metabolisme hormon seks.
TNF Alpha, IL-6
dan leptin yang secara kolektif disebut 'sitokin' dan terlibat dalam
mengirimkan pesan antara sel-sel.
Plasminogen
activator inhibitor-1 yang terlibat dalam pembekuan darah.
Angiotensin
yang terlibat dalam mengontrol tekanan darah.
Adiponektin
yang meningkatkan sensitivitas tubuh terhadap insulin sehingga membantu
melindungi terhadap diabetes tipe 2.
Lipoprotein
lipase dan apolipoprotein E yang terlibat dalam penyimpanan dan metabolisme
lemak untuk melepaskan energi.
Apa yang mungkin salah dengan jaringan adiposa?
Kedua jaringan adiposa terlalu banyak dan terlalu sedikit
dapat memiliki implikasi kesehatan yang parah. Lebih umum, jaringan adiposa
terlalu banyak menyebabkan obesitas, terutama dari terlalu banyak lemak
visceral. Obesitas menyebabkan sejumlah masalah kesehatan yang serius. Obesitas
meningkatkan risiko diabetes tipe 2 karena menyebabkan tubuh menjadi resisten
terhadap insulin. Hasil resistensi ini pada tingkat tinggi gula darah yang
buruk bagi kesehatan. Obesitas juga meningkatkan kemungkinan terjadinya tekanan
darah tinggi, kadar kolesterol tinggi dan peningkatan kecenderungan darah untuk
membeku. Semua ini meningkatkan risiko serangan jantung dan stroke.
Kurangnya jaringan adiposa (lipodistrofi) juga dapat
menyebabkan masalah yang sama dan terlihat dengan meningkatnya frekuensi
sebagai akibat dari obat yang digunakan untuk mengobati HIV / AIDS.
Pada gangguan makan (seperti anorexia nervosa), pasien
tidak makan makanan yang cukup untuk mempertahankan tingkat adiposa jaringan
mereka. Ini berarti bahwa mereka dapat kehilangan sejumlah berbahaya berat
badan.
UNTUK REFRENSI METABOLISME LEMAK LAINNYA KLIK INI
UNTUK REFRENSI METABOLISME LEMAK LAINNYA KLIK INI




{kind=link}